ÍNDICE
|
13. Tejido muscular 1. Muscular estriado 2. Muscular cardíaco 3. Muscular liso 14. Tejido nervioso 1. Neurona 2. Glia 15. Sangre 1. Composición de la sangre 2. Plasma sanguíneo 3. Células sanguíneas 4. Grupos sanguíneos 16. Resumen tejidos animales 17. Repaso 18. Prácticas 19. Otras presentaciones 20. Otras imágenes 21. Cuestiones |
2. ESQUEMAS


3. PRESENTACIONES




4. IMÁGENES DE TEJIDOS
Imágenes 1
Imágenes 3
Dibujos con los componentes
Tejidos
Tejidos 2
Tejidos 3
Tejidos animales 1
Tejidos animales 2
Tejidos animales 3
Tejidos animales 4
5. CONTENIDOS ANIMADOS

6. TEJIDOS ANIMALES
La histología es una disciplina eminentemente descriptiva basada en la observación mediante microscopios, tanto ópticos como electrónicos, de los distintos tipos de tejidos. Sin embargo, el conocimiento de la anatomía y organización de los tejidos es fundamental para comprender su fisiología y reconocer alteraciones patológicas, tanto de los propios tejidos como de los órganos y estructuras que forman.
Como se estudiar€ m€ás adelante, los cuatro principales tipos de tejidos aparecen precozmente en el periodo embrionario de desarrollo (en los dos primeros meses siguientes a la concepción). A partir de una “bola hueca” de c‚élulas, el blastocisto, se producen las
tres capas germinales primitivas denominadas ectodermo,endodermo y mesodermo gracias a una serie de movimientos en el blastocisto que se denominan gastrulación. A partir de estas tres capas germinales se formará€n los distintos tipos de tejidos en un proceso que se llama histiog‚énesis. A pesar de que las células que forman un organismo son muy diversas en forma y función, los histólogos han clasificado tradicionalmente a los tejidos animales en cuatro tipos fundamentales:
- Tejidos epiteliales. Conjunto de células estrechamente unidas que tapizan las superficies corporales, tanto internas como externas, y que además forman glándulas.
- Tejidos conectivos o conjuntivos. Agrupan a un variado tipo de tejidos que se caracterizan por la gran importancia de su matriz extracelular, la cuál en la mayoría de los casos es la principal responsable de su función. Se origina a partir de las células mesenquimáticas embrionarias. Forman la mayor parte del organismo y realizan funciones tan variadas como sostén, nutrición, reserva, etc. El tejido conectivo se especializa en diferentes tipos cuya clasificación puede depender del autor.
- Tejido muscular. Formado por células que permiten el movimiento de los animales gracias a la propiedad de sus células de contraerse.
- Tejido nervioso. Está constituido por células especializadas en procesar información. La reciben del medio interno o externo, la integran y producen una respuesta que envían a otras células.
|



CUESTIONES: 1 2 3 4 5 6
7. TEJIDO EPITELIAL
Los epitelios constituyen uno de los cuatro tejidos fundamentales de los animales. Están formados por células dispuestas de manera contigua, sin que exista prácticamente matriz extracelular, con lo que presentan una gran superficie de contacto entre ellas. En estas zonas adyacentes existen estructuras moleculares especializadas denominadas complejos de unión, como las uniones estrechas y desmosomas, que forman puentes intercelulares para fortalecer la cohesión entre las células epiteliales. Esto hace difícil o imposibilita el paso de determinadas moléculas por el espacio intercelular.
Los epitelios no poseen red de capilares sanguíneos por lo que la nutrición se realiza por difusión desde el tejido conectivo subyacente. Las células epiteliales se organizan formando uno o varios estratos que descansan sobre una capa de matriz extracelular especializada denominada lámina basal.
Bajo la lámina basal siempre aparece tejido conectivo. La lámina basal tiene un componente producido por las células epiteliales y otro por el tejido conectivo subyacente. Es característico también de los epitelios su polaridad, entendiendo por ello las diferencias morfofuncionales que presentan entre su dominio apical (orientado hacia la luz o hacia el exterior) y su dominio basal (orientado hacia la lámina basal).
Las funciones de los epitelios son muy variadas: protección frente a la desecación o la abrasión, filtración, absorción selectiva, transporte de sustancias por su superficie, y además pueden poseer células que actúan como órganos sensoriales, de secreción, etcétera. Algunas de estas funciones son posibles gracias a la presencia de especializaciones celulares en sus superficies libres o apicales como cilios, flagelos y microvellosidades.
El tejido epitelial recibe distintos nombres según donde se localize. Por ejemplo, en la piel se denomina epidermis, cuando recubre cavidades internas como la cavidad cardíaca, pulmonar o abdomen se llama mesotelio, y el epitelio que forma la superficie interna de los vasos sanguíneos y linfáticos es el endotelio. Además, los epitelios se nombran teniendo en cuenta el número de capas de células, la forma de las células de la capa más externa y si tienen o no especializaciones en su superficie apical.
Determinados epitelios pueden diferenciar y organizar sus células para formar estructuras macroscópicas especializadas como el pelo, las uñas o las plumas de las aves. Estas estructuras son inducidas por el tejido conectivo subyacente.
Las funciones de los epitelios son muy variadas: protección frente a la desecación o la abrasión, filtración, absorción selectiva, transporte de sustancias por su superficie, y además pueden poseer células que actúan como órganos sensoriales, de secreción, etcétera. Algunas de estas funciones son posibles gracias a la presencia de especializaciones celulares en sus superficies libres o apicales como cilios, flagelos y microvellosidades.
El tejido epitelial recibe distintos nombres según donde se localize. Por ejemplo, en la piel se denomina epidermis, cuando recubre cavidades internas como la cavidad cardíaca, pulmonar o abdomen se llama mesotelio, y el epitelio que forma la superficie interna de los vasos sanguíneos y linfáticos es el endotelio. Además, los epitelios se nombran teniendo en cuenta el número de capas de células, la forma de las células de la capa más externa y si tienen o no especializaciones en su superficie apical.
Determinados epitelios pueden diferenciar y organizar sus células para formar estructuras macroscópicas especializadas como el pelo, las uñas o las plumas de las aves. Estas estructuras son inducidas por el tejido conectivo subyacente.
El origen embrionario de los epitelios puede ser seguido hasta las tres hojas embrionarias formadas durante la gastrulación. Por ejemplo, el epitelio epidérmico procede del ectodermo, los que forman los capilares sanguíneos proceden del mesodermo y el epitelio digestivo del endodermo.
En algunas ocasiones las células epiteliales se agrupan y se especializan en la secreción de diversas sustancias. Hablamos entonces de epitelio glandular. El epitelio no glandular se denomina epitelio de revestimiento
En algunas ocasiones las células epiteliales se agrupan y se especializan en la secreción de diversas sustancias. Hablamos entonces de epitelio glandular. El epitelio no glandular se denomina epitelio de revestimiento


Los epitelios de revestimiento forman una capa que tapiza las superficies externas (piel, pulmones o aparato digestivo) e internas (vasos sanguíneos, linfáticos y pleuras). Cuando recubren cavidades serosas del organismo como las pleuras se denominan mesotelios, pero cuando recubren la parte interna de los vasos sanguíneos o linfáticos se llaman endotelios.
Los epitelios de revestimiento se caracterizan por poseer muy poca matriz extracelular y sus células están fuertemente unidas por complejos de unión. Poseen una alta tasa de renovación celular debido a la proliferación de las células progenitoras presentes y a una muerte celular continuada.
Pueden poseer especializaciones celulares que les permiten ser receptores sensoriales y, según los organimos, desarrollar estructuras complejas como pelos, plumas o escamas.
Se clasifican según el número de capas celulares que presentan y la forma celular de la capa más apical. En los epitelios simples, constituidos por una sola capa de células, todas las células contactan con la lámina basal y también forman la superficie libre del epitelio.
Se clasifican según el número de capas celulares que presentan y la forma celular de la capa más apical. En los epitelios simples, constituidos por una sola capa de células, todas las células contactan con la lámina basal y también forman la superficie libre del epitelio.
Las células pueden ser aplanadas, cúbicas (igual de anchas que de altas) o prismáticas (más altas que anchas). En los epitelios pseudoestratificados todas las células contactan con la lámina basal, pero no todas alcanzan la superficie libre del epitelio puesto que unas son más altas que otras. Éste es un epitelio simple con apariencia de estratificado.
Los epitelios estratificados poseen dos o más capas de células en las que sólo una de las capas contacta con la lámina basal, mientras la capa más superficial forma la superficie libre. Los epitelios estratificados se clasifican en planos, cúbicos o prismáticos, según la forma de las células del estrato que delimita el espacio libre del epitelio. Los epitelios de transición tienen más de una capa de células pero su aspecto cambia dependiendo del estado en que se encuentre el órgano que tapizan.


Se encuentra tapizando parte de las nefronas (que estudiaremos en el capítulo correspondiente al aparato excretor), recubriendo la cámara anterior del ojo, la pared de los alvéolos pulmonares y la cara interna del tímpano.
Existen dos tipos especiales de epitelios planos monoestratificados:






Algunos epitelios de este tipo presentan cilios en la zona apical, como el epitelio que recubre el interior de la vesícula biliar o los oviductos.




El estrato córneo es la capa superficial. Las cé‚lulas en este estrato ya no poseen ni n†cleo ni orgánulos. La membrana plasm€ática estဠengrosada y cubierta en su superficie externa por glucolƒípidos; el interior de las células esta completamente lleno de queratina.






FUNCIONES DE LOS EPITELIOS DE REVESTIMIENTO

Estratificado
7.2. Epitelios glandulares
Las glándulas son asociaciones de células epiteliales altamente diferenciadas, que segregan sus productos de síntesis, bien a una superficie libre, mediante un conducto secretor (glándulas exocrinas), o bien directamente al torrente sanguíneo (glándulas endocrinas o de secreción interna).
Todas las células del organismo del organismo están dotadas de actividad secretora. La pro- piedad de secreción sólo se convierte en función cuando moviliza total o casi totalmente la actividad de la célula y cuando conduce a la elaboración de productos utilizados por el orga- nismo. Una célula capacitada para tal función tiene una actividad glandular
La secreción se desarrolla según un proceso constante cualquiera que sea el tipo celular. Comprende una sucesión de cuatro fases:
 Las glándulas se forman a partir de superficies epiteliales. El epitelio prolifera y se hunde, en el tejido conjuntivo, manteniendo, o no, contacto con la superficie, según que vaya a ser glándula exocrina o endocrina, respectivamente. En las glándulas endocrinas, las células pueden agruparse en cordones o en folículos. En los folículos, el producto de secreción se acumula en gran cantidad.
Las glándulas se forman a partir de superficies epiteliales. El epitelio prolifera y se hunde, en el tejido conjuntivo, manteniendo, o no, contacto con la superficie, según que vaya a ser glándula exocrina o endocrina, respectivamente. En las glándulas endocrinas, las células pueden agruparse en cordones o en folículos. En los folículos, el producto de secreción se acumula en gran cantidad.

En las Glándulas exocrinas el producto se vierte al medio externo bien por un conducto excretor, bien directamente. El medio externo puede ser:


Según el producto de secreciÅn las glándulas se suelen dividir en MUCOSAS si vierten mucopolisacáridos, como por ejemplo las existentes en la mucosa intestinal y SEROSAS si su producto es fundamentalmente proteico, en concreto enzimático. En algunos casos las glándulas fabrican lípidos como las glándulas sebáceas de la piel o lipoproteínas, como las glándulas mamarias que fabrican leche.
Según la forma de secreción las glándulas se clasifican en:
Merocrinas: que es una forma de secreción que respeta la integridad de la células, es decir, ésta no muere ni sale dañada en el proceso. la mayoría de las glándulas pertenecen a este tipo (salivares, páncreas, etc.)
Apocrinas: es una forma de secreción en la cual el producto de secreción acumulado en el polo apical de la célula se expulsa con éste. Las células así decapitadas regeneran rápidamente esta pérdida de sustancia y comienzan nuevamente un ciclo secretor.
sustancia y comienzan nuevamente un ciclo secretor.
Son ejemplos típicos de glándulas apocrinas, las glándulas mamarias, algunas glándulas su- doríparas del hueco axilar, de los pliegues inguinales, de los órganos genitales, de la región perianal y de los párpados (las llamadas glándulas de Moll)

 Holocrinas: tipo de secreción en la que la cesión de sustancias lleva consigo la muerte celular. En este caso, la célula está completamente llena de se- creción. La secreción holocrina tiene lugar, sobre todo, en las glándulas sebáceas de la piel.
Holocrinas: tipo de secreción en la que la cesión de sustancias lleva consigo la muerte celular. En este caso, la célula está completamente llena de se- creción. La secreción holocrina tiene lugar, sobre todo, en las glándulas sebáceas de la piel.

Las glándulas también pueden clasificarse atendiendo al número de células y disposición de
Glándulas unicelulares: la glándula es una sola célula. Como ejemplo cita- remos a las ya mencionadas glándulas o células caliciformes. Estas células fabricantes de moco y glucoproteínas tienen forma de copa y se alternan entre las células de un epitelio de revestimiento. Vierten su contenido al exterior para formar una capa protectora.
Glándulas pluricelulares: se dividen en simples, ramificadas y compuestas. En las glándulas ramificadas, varias unidades secretoras individuales desembocan en un conducto excretor común. En las glándulas compuestas, el conducto secretor principal está subdividido en varios conductos más pequeños.
Según el aspecto de la unidades glandulares secretoras, también llamadas unidades glandulares , las glándulas se clasifican también como acinosas (en forma de acino, como frutos en baya), alveolares (en forma de sáculos) y tubulares (en forma alargada, como un tubo). Existen formas mixtas que son glándulas tubuloacinosas y las tubuloalveolares.
Las unidades glandulares están formadas por células secretoras rodeadas por una lámina basal.

7.3. Epitelios sensoriales
Los epitelios sensoriales, altamente especializados, intervienen en la captación de impresiones sensoriales, serán tratados en el tema dedicado a los sentidos.
8. TEJIDOS CONECTIVOS
 Existen varios tipos de tejidos conjuntivos localizados en diversos sitios del organismo, adaptados a funciones específicas tales como:
Existen varios tipos de tejidos conjuntivos localizados en diversos sitios del organismo, adaptados a funciones específicas tales como:


Células mesenquimatosas: Son células procedentes del mesodermo presentes en el mesénquima o tejido conjuntivo embrionario. Se puede decir que constituyen un tipo de fibroblasto primitivo o su precursor. Apare- cen también en el tejido mucoide del cordón umbilical. Estas células pueden diferenciarse hacia fibroblastos como ya hemos dicho, adipocitos, células reticulares, miofibroblastos, células musculares lisas o hasta células de Leydig.
Células reticulares: Únicamente se hallan presentes en el tejido conjuntivo reticular que forma parte del estroma de los órganos linfoides y hematopoyéticos.
Reciben este nombre por sus prolongaciones estrelladas por las que se unen a otras células
gracias a desmosomas.
Fibroblastos: Son las células características del tejido conjuntivo y están presentes en la mayor parte de sus variedades y, sobre todo, en las que se encuentran en pleno desarrollo. Se distribuyen en el tejido conjuntivo entre las fibras de colágeno que ellos mismos fabrican. También fabrican la sustancia fundamental.
En general, los fibroblastos tienen forma irregu- lar: son células aplanadas y fusiformes, provis- tas de prolongaciones delgadas que entran temporalmente en contacto con las de los fibro- blastos contiguos. Existen dos tipos de fibroblas- tos y todas las fórmulas intermedias:
El fibroblasto típico o fibroblasto jo- ven, con una intensa actividad de síntesis. Éste tiene capacidad de dividirse si es conveniente- mente estimulado, por ejemplo cuando hay ne- cesidad de cicatrización.
El fibroblasto maduro o fibrocito, es poco activo. El fibroblasto tiene un núcleo de gran tamaño, ovoide y con un nucleolo voluminoso. El citoplasma contiene:
 Otras células del sistema de defensa:
Otras células del sistema de defensa:
- Monocitos que se transformarán en macrófagos.
- Linfocitos especialmente células plasmáticas.
- Leucocitos neutrófilos polimorfonucleares.
- Leucocitos eosinófilos.
Unas células particulares, Los adipocitos: Son células conjuntivas que acumulan, en forma de reserva, importantes cantidades de lípidos. Son las células típicas del TEJIDO ADIPOSO y existen dos tipos:
El tejido conectivo I: La circulación del agua.
El tejido conectivo II: Las fibras.
El tejido conectivo III: Las células.
El tejido conectivo IV: La síntesis de fibras colágenas.
Lámina 6. Fibras del tejido conectivo.
Lámina 7.- Células del tejido conectivo.
Conectivo típico
Laxo: (esófago, estómago, tráquea, arteria, vena)
Regular: (tendón)
9. TEJIDOS CONJUNTIVOS
Las funciones de este tejido son:



9.3. TEJIDO CONJUNTIVO FIBROSO DENSO
Este tipo de tejido se caracteriza por la abundancia de fibras de colágeno y de elastina, y por la escasez de células. Los fascículos de fibras de colágeno siguen trayectos que dependen de la dirección de las fuerzas que tiene que soportar el órgano en cuestión. Se encuentran dos tipos de tejido:
A) TEJIDO CONJUNTIVO FIBROSO DENSO NO ALINEADO
En él, las fibras de colágeno no están orientadas y se distribuyen de manera compacta. Estos tejidos están sometidos a fuerzas multidireccionales. Se encuentran en la duramadre (una meninge = membrana que hay entre el cráneo y el encéfalo), en las articulaciones, en la esclerótica del ojo, formando parte de la cápsula (parte más externa de un órgano y que le da la forma) del testículo, bazo, riñón, hígado y otros. También forma la vaina de los tendones ( un tendón es un cordón fibroso de color blanco nacarado mediante el cuál el músculo se une al hueso o a otro órgano) y el periostio.



TEJIDO CONJUNTIVO FIBROSO DENSO ALINEADO
Sus fibras de colágeno se encuentran todas alineadas en el sentido de la tracción. Los fibroblastos de este tejido reciben el nombre de células tendinosas. Este tejido está adaptado para transmitir fuerzas de tracción mecánica: en efecto, es poco extensible (5%) y muy resistente (100 kg por cm2 de sección). Se localiza en los tendones (que transmiten a los
huesos el efecto de tracción desarrollados por los músculos) y en los ligamentos (un ligamento es una cinta o fascículo que se inserta en huesos y cartílagos y que sirve para como medio de unión de la articulaciones o para otros fines). Es también un tejido típico de la córnea.



9.4. TEJIDO CONJUNTIVO ELÁSTICO
Es un tejido rico en fibras elásticas aunque también hay de colágeno. Las fibras pueden estar irregularmente distribuidas, pero también pueden disponerse muy ordenadas: en haces de fibras paralelas, rodeados y separados entre sí, por un conjuntivo de fibras de reticulina.
Se encuentra formando los ligamientos amarillos de los mamíferos. En el hombre además en las cuerdas vocales, el ligamiento estilohiodeo en la laringe, el ligamento suspensor del pene y en los tendones del músculo de la tráquea. También se encuentra en la túnica media de las arterias elásticas.


10. TEJIDO ADIPOSO COMÚN
En la especie humana el tejido adiposo constituye aproximadamente un 10% del peso corporal, y su distribución varía del sexo masculino al femenino. Así en los hombres es abundante en la nuca, en la 7ª vértebra cervical, sobre el deltoides y el tríceps. En mujeres, en los pechos, nalgas y cara anterior del muslo.
Se puede considerar como una variante de tejido conjuntivo altamente especializada en el almacenamiento de lípidos. En los mamíferos se encuentra en muchos lugares en donde hay tejido conjuntivo laxo, pero forma acumulaciones mayores en la hipodermis, mesenterios (son distintos repliegues del peritoneo que fijan las distintas porciones del intestino a las paredes abdominales). Este tejido tiene un color blanco-amarillento debido al contenido de carotenoides.



Metabolismo del tejido adiposo
Se basa en el almacenamiento y liberación de triglicéridos según las siguientes etapas:
Constituye una variante del tejido adiposo, que se presenta en embriones de mamíferos, siendo más raro en los adultos.
Los adipocitos de grasa parda son células más pequeñas que las del tejido adiposo común. El núcleo no está rechazado pudiendo ser incluso central. El citoplasma no contiene una sola gota de grasa sino muchas gotitas de lípidos de tamaño variable. Se trata de una estructura microvacuolar.
El color de este tejido se debe a la abundancia de citocromos que tienen las mitocondrias de las células de este tejido.
El adipocito blanco.
El adipocito pardo.
Lámina 8.- Tejido adiposos.
Adiposo: (tráquea)
11. TEJIDO CARTILAGINOSO
Es, junto con el hueso, uno de los principales tejidos de soporte de los animales. Su función es posible gracias a las propiedades de su matriz extracelular, la cual es predominante en este tipo de tejido. El cartílago es una estructura semirígida que permite mantener la forma de numerosos órganos, recubre la superficie de los huesos en las artículaciones y es el principal tejido de soporte durante el desarrollo embrionario, cuando el hueso aun no está formado. Es un tejido avascular y su matriz extracelular está formada fundamentalmente por colágeno, fibras elásticas y glucosaminoglicanos sulfatados. Las células que lo componen son los condrocitos, los cuales se localizan en pequeñas oquedades diseminadas denominadas lagunas.
La mayor parte del cartílago, excepto el tipo de cartílago denominado fribrocartílago, está rodeado por una capa de tejido conectivo denominada pericondrio. Éste posee una capa externa, denominada pericondrio fibroso, formada por tejido conectivo fibroso que contiene fibroblastos y fibras de colágeno, y otra interna denominada pericondrio condrogénico, donde se encuentran las células condrogénicas y los condroblastos que por diferenciación darán lugar a los condrocitos.
Hay tres tipos de cartílago en el organismo: hialino, elástico y fibrocartílago.
11.1. Cartílago hialino
Tejido de aspecto cristalino, blanco azulado. Lo encontramos en: caras articulares de los huesos, cartílagos costales, cartílagos de nariz, laringe y traquea, en el feto casi todo el esqueleto es cartílago hialino. Los condrocitos ocupan unas pequeñas cavidades dentro de la matriz rodeados de una laguna.
Formado por células esféricas, nucleo grande y centrado con uno o más nucleolos, citoplasma basófilo, grandes mitocondrias y vacuolas con gotas de agua y glucógeno. En el cartilago vivo los condrocitos ocupan la totalidad de las lagunas, después de un proceso de deshidratación y fijación se produce una cierta concentración de los condrocitos y las células pueden presentar una distorsión notable.



11.2. Cartílago fibroso
Es un carílago intermedio entre el cartílago hialino y el tejido conjuntivo denso alineado. Se caracteriza por su riqueza en fibras de colágeno de diverso grosor, que le confiere gran resis- tencia a las tracciones y compresiones. El colágeno enmascara a toda la sustancia fundamental excepto en la proximidad de los condrocitos.
Esta variedad de cartílago carece de pericondrio.
Se encuentra en los discos intervertebrales, en la sínfisis pélbica, en el menisco y en los puntos en los que los tendones y ligamentos se insertan en el hueso.



El tejido cartilaginosos.
Lamina 11.- Tejido cartilaginoso.
Cartilaginoso Hialino: (tráquea)
12. TEJIDO ÓSEO
El tejido óseo, al igual que el cartilaginoso, es un tejido de sostén en el que la matriz rígida está impregnada por un complejo proteinocálcico que envuelve a las células óseas u osteocitos y sostenida por un armazón de colágeno.
Este tejido cumple varias funciones:
Sustancia intercelular mineral: La sustancia intercelular de hueso es dura y presenta unos huecos intercomunicados entre sí en donde se alojan las células óseas.
Las sales minerales del tejido óseo son esencialmente:
- Osteoblastos
- Osteocitos
- Osteoclastos



12.1. CLASIFICACIÓN DEL TEJIDO ÓSEO


Huesos planos: como el parietal, frontal, omoplato, ... Están cubiertos externamente por tejido óseo compacto y en el interior contienen tejido óseo esponjoso.
Huesos cortos: como los huesos de la falanges. Están formados por tejido óseo esponjoso rodeado de periostio.

El tejido óseo I: las células y sustancia intercelular.
El tejido óseo II: el hueso haversiano.
La osificación endocondral I: el modelo cartilaginoso.
La osificación endocondral II: el manguito óseo.
La osificación endocondral III: la calcificación del cartílago.
La osificación endocondral III: la remodelación.
La articulación sinovial. .
Lámina 12.- Tejido oseo.
Lamina 13.- Un hueso
Compacto: (hueso compacto)
Esponjoso: (hueso esponjoso)
13. TEJIDO MUSCULAR
Este tejido, de origen mesenquimático, está constituído por: células musculares (fibras musculares), capaces de generar movimientos al contraerse bajo estímulos adecuados y luego relajarse y tejido conjuntivo estrechamente asociado a las células musculares. Este actúa como sistema de amarre y acopla la tracción de las células musculares para que puedan actuar en conjunto. Además, conduce los vasos sanguíneos y la inervación propia de las fibras musculares.
Tipos de tejido muscular
En los vertebrados, se distinguen 3 tipos de músculo:


13.1. Tejido muscular esquelético
Este tejido está formado por manojos de células cilíndricas muy largas (de hasta 30 cm), multinucleadas y estriadas transversalmente, llamadas también fibras musculares esqueléticas. Los núcleos de las fibras se ubican vecinos a la membrana plasmática (sarcolema), que aparece delimitada por una lámina basal (lámina externa). El tejido conjuntivo que rodea a las fibras musculares contiene numerosos vasos sanguíneos y nervios y se dispone de manera de provocar la contracción de las fibras musculares a los sitios de inserción del músculo.
Cada fibra muscular recibe una terminación del axón de una neurona motora, formándose en la zona de unión una estructura denominada placa motora.
El músculo esquelético se une a los huesos a través de los tendones, estructuras continuas con la envoltura conjuntiva llamada epimisio, que rodea externamente al músculo completo. El tejido conjuntivo penetra al interior del músculo formando el perimisio, que corresponde a delgados septos de tejido conjuntivo que envuelven a manojos o fascículos de fibras musculares. A partir del perimisio, se origina el endomisio formado por delgadas vainas de fibras reticulares que rodean cada una de las fibras musculares. Los vasos sanguíneos penetran al músculo a través de estos septos conjuntivos.
La inervación del tejido muscular esquelético se relaciona directamente con la regulación de la contracción de cada fibra muscular y en consecuencia con el estado de tensión del músculo completo.


Fibras musculares esqueléticas
Las fibras musculares esqueléticas se caracterizan por la presencia de estriaciones transversales periódicas. Esta estriación resulta de la existencia en su citoplasma de las miofibrillas estructuras responsables de la contracción muscular.
Las miofibrillas son estructuras cilíndricas largas (1 a 3 mm de diámetro) que corren paralelas al eje logitudinal de la célula, y están formados por miofilamentos finos y miofilamentos gruesos, dispuestos en tal forma que inducen la apariencia de bandas claras y oscuras que se repiten a a lo largo de cada miofibrilla, determinando la organización de los sarcómeros.
La banda oscura se conoce como banda A (de anisótropa) y la clara como banda I (de isótropa). Cada banda I aparece bisectada por una línea transversal oscura denominada disco o línea Z . Al centro de la banda A hay una zona más clara que corresponde a la banda H en cuyo centro está la línea M.
En el citoplasma que rodea a las miofibrillas se disponen tanto las miocondrias como las cisternas del retículo sarcoplásmico a las cuales se asocian los túbulos T, en una organizacion precisa con respecto a los sarcómeros y repetidas a todo lo largo de las células musculares esqueléticas .


Se distinguen 3 tipos de fibras musculares esqueléticas: rojas, blancas e intermedias.
Las fibras rojas, que abundan en los músculos rojos, son de diámetro pequeño y contienen gran cantidad de mioglobina y numerosas mitocondrias, que se disponen en filas entre las miofibrillas y en acúmulos por debajo del sarcolema. Los músculos rojos se contraen más lentamente, por lo que se ha asumido que la fibra roja es una fibra lenta.
Las fibras blancas, presentes en los músculos blancos, son de diámetro mayor, poseen menor cantidad de mioglobina y un número menor de mitocondrias que se disponen, de preferencia, entre las miofibrillas, a nivel de la banda I. En este tipo de fibras la línea Z es mas delgada que en las fibras rojas.
Las fibras intermedias presentan características intermedias entre las otras 2 variedades de fibras, pero superficialmente se asemejan más a las fibras rojas y son mas abundantes en los músculos rojos. Poseen un número de mitocondrias equivalente al de las fibras rojas, pero su línea Z es delgada como en las fibras blancas.
Regulación de la contracción del sarcómero
La contracción muscular está regulada por variaciones en los niveles citosólicos de Ca++, los que afectan las interacciones entre las cabezas de miosina y los filamentos de actina a través de las 2 proteínas accesorias asociadas a actina en el filamento fino: tropomiosina y troponina.
En el músculo en reposo (concentración citosólica de la miosina no puede asociarse a la actina debido a que los sitios de unión para las cabezas de miosina en las G-actina, están bloqueados por la tropomiosina. Al aumentar las concentraciones citosólicas de Ca, la subunidad TnC de la troponina une Ca, produciéndose un cambio conformacional de la molécula de troponina y el desplazamiento de la molécula de tropomiosina hacia la parte mas profunda de la hendidura de la hélice de actina. Como consecuencia los sitios, en la G-actina, capaces de interactuar con las cabezas de la miosina quedan libres.
Las variaciones en las concentraciones citosólicas de Ca se producen en respuesta a los estímulos nerviosos que inducen la contracción muscular y que actúan desencadenando la liberación de Ca desde el retículo sarcoplásmico hacia el citosol.
Estructura de las miofibrillas
La unidad estructural y funcional de las células musculares estriadas es el sarcómero, elemento que se repite a lo largo de las miofibrillas. El análisis de la estructura y composición molecular del sarcómero, permite entender el mecanismo de contracción de las fibras musculares estriadas, basado en el deslizamiento de los miofilamentos gruesos sobre los miofilamentos finos.
Los filamentos gruesos (de 15 nm de ancho y 1.6 mm de largo) están formados principalmente por miosina y se localizan a lo largo de la banda A. Los filamentos finos (de 8 nm de ancho y 1.0 mm de largo) corresponden a microfilamentos de F-actina. Estos anclan en la línea Z, luego cursan a lo largo de la banda I y penetran la banda A, donde corren paralelos a los filamentos gruesos, terminando a nivel de la banda H que contiene sólo filamentos gruesos. En la banda A se observan puentes que se extienden desde los filamentos gruesos hacia los filamentos finos y que corresponden a las cabezas de las moléculas de miosina. A nivel de la línea M cada filamento grueso se asocia a 6 filamentos gruesos adyacentes, a través de puentes proteicos dispuestos radialmente.
Durante el proceso de contracción, los filamentos finos de los sarcómeros adyacentes son empujados hacia el centro de la banda A, lo que produce el acortamiento del sarcómero. Como consecuencia de este proceso, se oblitera la banda H y disminuye la longitud de la banda I, sin que se modifique la longitud de la banda A. El grado de traslapamiento entre filamentos gruesos y finos explica este fenómeno.



Los filamentos finos están formados por actina, tropomiosina y troponinas, proteínas que se relacionan directamente con el proceso de acortamiento del sarcómero .
Los microfilamentos de actina están constituídos por 2 hebras proteicas, que se enrollan para formar una estructura helicoidal doble. Cada hebra corresponde a un polímero de moléculas asimétricas de G actina, lo que otorga a los microfilamentos de actina una polaridad definida.
La molécula de miosina-II está formada por 2 polipéptidos (cadenas pesadas ) y 4 polipéptidos (cadenas livianas). Está organizada en 3 dominios estructural y funcionalmente distintos: cabeza, cuello y cola. En el extremo NH2-terminal las 2 cadenas pesadas presentan una estructura globular, llamada cabeza, la que se continúa en una zona con forma de bastón, de unos 150 nm de largo, cuya porción inicial corresponde al cuello de la molécula y el resto a la cola.
Retículo Sarcoplásmico y túbulos T (túbulos transversos)
En el músculo esquelético, cada miofibrilla está rodeada de un elaborado sistema de membranas lisas que corresponden al retículo sarcoplásmico. Estas membranas están alineadas en forma precisa con respecto al patrón de bandeo de las miofibrillas. En la zona de unión de la banda A con la banda I el retículo sarcoplásmico se expande para formar las cisternas terminales. Las 2 cisternas terminales paralelas se asocian estrechamente a un tubo transverso (T), formando un complejo denominado tríada .
La contracción de una fibra muscular requiere de la contracción simultánea de todas sus miofibrillas. La forma y distribución del sistema T permite que la onda de depolarización, responsable de la contracción muscular, se distribuya rápidamente desde la superficie celular hacia el interior del citoplasma alcanzando a cada miofibrilla.
La depolarización de la membrana plasmática de la célula muscular, que se propaga a lo largo de los túbulos T, produce la apertura de canales de Ca++ en la membrana del retículo sarcoplásmico y la liberación de Ca++ hacia el citosol. Se piensa que la onda de depolarización induce un cambio conformacional en proteínas sensoras del túbulo T, que se transmite directamente a la proteína que forma los canales de Ca++ del retículo sarcoplásmico.


Placa motora (unión neuromuscular)
La membrana plasmática de la célula muscular está electricamente polarizada. Un estímulo apropiado depolariza la membrana y produce la contracción. Normalmente, la depolarización es producida por un estímulo nervioso. El músculo está inervado por terminales nerviosas de neuronas motoras de la médula espinal. En el punto de inervación, el nervio pierde su vaina de mielina, y se asocia a una región especializada de la superficie de la fibra muscular, para formar la placa motora. En una fibra muscular esquelética, cada terminal axónico motor forma sólo una placa motora.
En la zona de contacto, la terminal axónica forma una dilatación que se aloja en una depresión poco profunda de la superficie de la fibra llamada hendidura sináptica primaria .
En la zona de unión, la lámina basal de la célula de Schwann se fusiona con la lámina basal de la célula muscular. Esta cubierta celular fusionada se extiende hacia la hendidura sináptica primaria, separando la fibra nerviosa de la fibra muscular, y penetra al interior de cada hendidura sináptica secundaria. Cuando el potencial de acción alcanza a la placa motora, el neurotransmisor acetilcolina, contenido en las vesículas sinápticas, se libera y difunde a través de la hendidura. Este mediador se une a receptores de acetilcolina presentes en la membrana postsináptica, concentrados principalmente en la entrada de los pliegues sinápticos secundarios, e induce la depolarización del sarcolema que es transmitida a los tubos T.


Unidad motora: conjunto de fibras musculares esqueléticas inervadas por ramificaciones del axón de una misma neurona motora y que, en consecuencia, son estimuladas simultáneamente a contraerse.
Ramas de una misma motoneurona pueden llegar a inervar hasta 500 fibras musculares. Sin embargo, mientras más fino el movimiento que debe efectuar el músculo, menor es el tamaño de la unidad motora, existiendo situaciones en que cada fibra nerviosa inerva sólo una fibra muscular.
El tejido muscular estriado I: organización.
El tejido muscular estriado II.- el citoesqueleto
El tejido muscular estriado III:.el aparato contráctil. la contracción muscular
El tejido muscular estriado IV: La fibra muscular esquelética.
El tejido muscular estriado V: La fibra roja.
El tejido muscular estriado VI: La fibra blanca.
El musculo esquelético: organización.
Lámina 15- Músculo estriado.
Lámina 16- Organización del músculo
Fibra estriada: (esófago, lengua)
13.2. Músculo cardíaco
El músculo cardíaco está formado por células musculares ramificadas, de estructura estriada y contacción involuntaria, que poseen 1 o 2 núcleos y que se unen entre sí a través de un tipo de unión propia del músculo cardíaco llamada disco intercalar. A diferencia del músculo esquelético, las fibras musculares cardíacas corresponden a un conjunto de células cardíacas unidas entre sí en disposición lineal.
Las células musculares cardíacas, de unos 15 m m de diámetro y unos 100mm de largo, tienen el núcleo ubicado al centro del citoplasma y presentan estriaciones transversales similares a las del músculo esquelético. El retículo sarcoplásmico no es muy desarrollado y se distribuye irregularmente entre las miofibrillas, que no aparecen claramente separadas. Sin embargo, las mitocondrias, que son extremadamente numerosas, están distribuidas regularmente dividiendo a las células cardíacas en miofibrillas aparentes. En el sarcoplasma hay numerosas gotas de lípido y partículas de glicógeno. Las células están rodeadas por una lámina externa, comparable a la lámina basal de los epitelios.



El musculo cardiaco: la fibra muscular común.
El musculo cardiaco: la fibra de Purkinje.
Cardiaco:(músculo cardiaco)
13.3. Músculo Liso
El músculo liso está formado por fibras musculares lisas que corresponden a células uninucleadas, delgadas y aguzadas en los extremos, cuya longitud varía entre 20 y 500 mm. Este tipo de músculo forma la porción contráctil de la pared de diversos órganos tales como tubo digestivo y vasos sanguíneos, que requieren de una contracción lenta y sostenida. Las células se organizan en grupos, formando haces, rodeados de tejido conjuntivo fibroso que contiene vasos sanguíneos.
El núcleo de las fibras musculares lisas se ubica en el centro de la fibra y los organelos citoplasmáticos tales como mitocondrias, aparato de Golgi, retículo endoplásmico rugoso y ribosomas libres se localizan, mayoritariamente, en la vecindad de los polos nucleares. El resto del citoplasma está ocupado por abundantes miofilamentos finos de actina, una proporción menor de miofilamentos gruesos de miosina, y un citoesqueleto de filamentos intermedios formados por desmina.


Las fibras musculares lisas se disponen desplazadas una respecto de la otra, de manera que el extremo delgado de una fibra se ubica vecino a la parte ancha de la fibra vecina. Esta disposición de las fibras y la localización del núcleo en el centro, explica el aspecto del músculo liso en corte transversal .
Las fibras musculares lisas están rodeadas por una lámina basal (lámina externa) comparable a la lámina basal de los epitelios
El aparato contráctil del músculo liso se contrae mas lentamente que el del músculo estriado, pero permite un acortamiento mayor de las fibras musculares lisas. El mecanismo de contracción, en esta variedad de músculo, también se basa en el deslizamiento de los filamentos finos sobre los filamentos gruesos. Los filamentos de actina de las fibras musculares lisas son fáciles de detectar a nivel ultraestructural; en cambio la visualización de los filamentos gruesos requiere de condiciones de fijación especiales, que demuestran que en el músculo liso por cada filamento grueso hay una proporción mucho mayor de filamentos finos (1:14) que la que se observa en el músculo esquelético (1:6).
En estas células, la contracción es regulada también por alza en las concentraciones citosólicas de Ca++. Sin embargo, la regulación de la contracción está asociada a miosina y no a actina. Un alza en las concentraciones citosólicas de Ca++ induce la fosforilación de las cadenas livianas de la miosina lo que produce una modificación en la cola de la molécula que permite la formación de filamentos gruesos y genera un cambio conformacional en la cabeza que permite su interacción con actina.
Inervación del Músculo Liso
El músculo liso está inervado por nervios de los sistemas simpático y parasimpático. Con frecuencia, los axones de los nervios terminan en una serie de dilataciones en el conjuntivo que rodea a las células musculares. Algunas de estas dilataciones axónicas están muy próximas (10-20 nm) a la superficie de la célula muscular dando origen a uniones neuromusculares. De acuerdo a la proporción de células inervadas en un determinado músculo, se distinguen:
La capacidad de regeneración es diferente en las 3 variedades de músculo.
El músculo esquelético tiene la capacidad de regenerar parcialmente a partir de las llamadas células satélite. Estas corresponden a células uninucleadas, fusiformes que yacen dentro de la lámina basal que rodea a cada fibra. Se considera que corresponden a mioblastos que persisten luego de la diferenciación del músculo. Frente a daño muscular u otros estímulos estas células, relativamente escasas, se activan, proliferan y se fusionan para formar nuevas fibras.
El músculo cardíaco no tiene, practicamente, capacidad de regenerar. Los daños del músculo cardíaco se reparan por proliferación del tejido conjuntivo, produciéndose una cicatriz.
El músculo liso tiene tambien una capacidad de regeneración moderada. Luego de daño muscular, algunas células musculares lisas entran en mitosis y reemplazan el tejido dañado. Si la capacidad de proliferación no es suficiente para reparar el daño, se produce una cicatriz de tejido conjuntivo.

ANIMACIONES



Es el tejido propio del Sistema Nervioso el cuál, mediante la acción coordinada de redes de células nerviosas:

El Sistema Nervioso Central (SNC), se origina desde el epitelio del tubo neural y su tejido nervioso contiene neuronas, células de neuroglia y capilares.
El Sistema Nervioso Periférico (SNP), que conecta los receptores sensoriales con SNC. y a este con las células efectoras, se desarrolla a partir de la cresta neural y sus células se asocian a otros tejidos del organismo. Sin embargo, es una extensión del tejido nervioso del SNC ya que zonas de las neuronas sensitivas y efectoras y todas las interneuronas se encuentran en el SNC, mientras que los ganglios nerviosos y los nervios periféricos corresponde al tejido nervioso propio del SNP
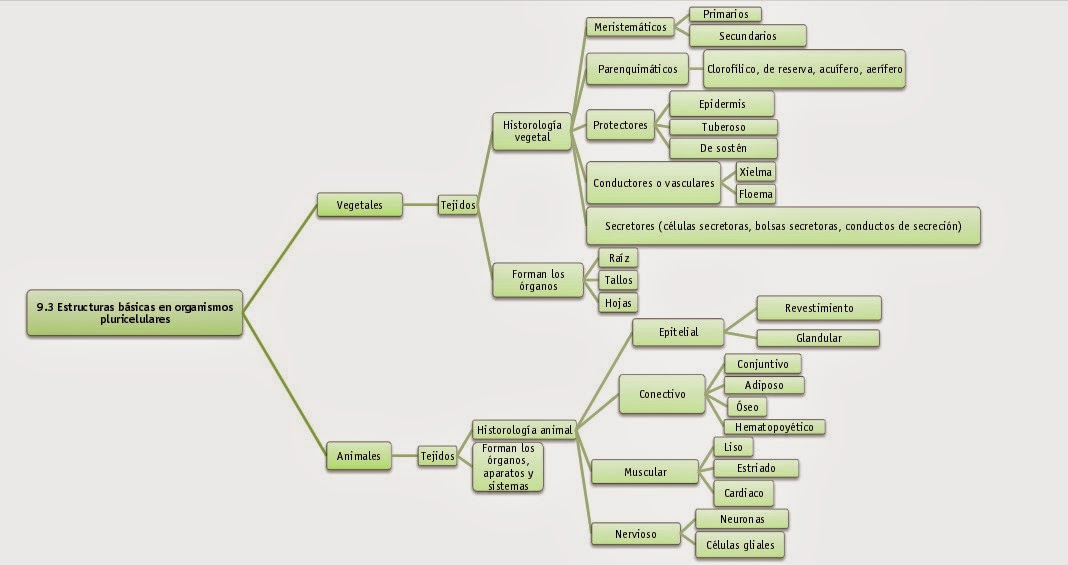
11.1. Neuronas
Son las células funcionales del tejido nervioso. Ellas se interconectan formando redes de comunicación que transmiten señales por zonas definidas del sistema nervioso . Los funciones complejas del sistema nervioso son consecuencia de la interacción entre redes de neuronas, y no el resultado de las características específicas de cada neurona individual.
La forma y estructura de cada neurona se relaciona con su función específica, la que puede se:


Cada zona de las células nerviosas se localiza de preferencia en zonas especializadas del tejido nervioso:
Según el número y la distribución de sus prolongaciones, las neuronas se clasifican en:
Bipolares, que además del axón tienen sólo una dendrita; se las encuentra asociadas a receptores en la retina y en la mucosa olfatoria
Speudo-unipolares, desde las que nace sólo una prolongación que se bifurca y se comporta funcionalmente cono un axón salvo en sus extremos ramificados en que la rama periférica reciben señales y funcionan como dendritas y transmiten el impulso sin que este pase por el soma neuronal; es el caso de las neuronas sensitivas espinales
Multipolares desde las que, además del axón, nacen desde dos a más de mil dendritas lo que les permite recibir terminales axónicos desde múltiples neuronas distintas. La mayoría de las neuronas son de este tipo. Un caso extremo do lo constituye la célula de Purkinje que recibe más de 200.000 terminales nerviosos

Las neuronas son células sintetizadoras de proteínas, con un alto gasto de energía metabólica, ya que se caracterizan por:
Las principales funciones del axón son:
En el SNC los axones están rodeados por la mielina de los oligodendrocitos (fibras nerviosas mielínicas del SNC), mientras que en el SNP pueden estar rodeados, ya sea, por prolongaciones citoplasmáticas de las células de Schwann (fibras amielínicas) o por la mielina las células se Schwann (fibras nerviosas mielínicas del SNP)
Los impulsos nerviosos son ondas transitorias de inversión del voltaje que existe a nivel de la membrana celular, que se inician el sitio en que se aplica el estímulo. Cada una de estas ondas corresponde a un potencial de acción. Este proceso es posible porque entre las macromoléculas que, como proteínas integrales, ocupan todo el espesor del axolema se encuentran:
Además, en el interior de la neurona existen proteínas e iones con carga negativa. Esta diferencia de concentración de iones produce también una diferencia de potencial entre el exterior de la membrana y el interior celular. El valor que se alcanza es de unos -70 milivoltios (negativo el interior con respecto al valor de cargas positivas del exterior).
Esta variación entre el exterior y el interior se alcanza por el funcionamiento de la bomba de sodio/potasio (Na+/K+)
La bomba de Na+/K+ gasta ATP. Expulsa tres iones de sodio que se encontraban en el interior de la neurona e introduce dos iones de potasio que se encontraban en el exterior. Los iones sodio no pueden volver a entrar en la neurona, debido a que la membrana es impermeable al sodio. Por ello, la concentración de iones sodio en el exterior es elevada. Además, se pierden 3 cargas positivas cada vez que funciona la bomba de Na+/K+, aunque entren dos cargas de potasio. Esto hace que en el exterior haya más cargas positivas que en el interior, creando una diferencia de potencial. Se dice que la neurona se encuentra en potencial de reposo, dispuesta a recibir un impulso nervioso.


Cuando el impulso nervioso llega a una neurona en estado de reposo la membrana se despolariza, abriéndose los canales para el sodio. Como la concentración de sodio es muy elevada en el exterior, cuando los canales para el sodio se abren se invierte la polaridad, con lo que el interior de la neurona alcanza un valor electropositivo, respecto del exterior.
Si la despolarización provoca un cambio de potencial de 120 milivoltios más de los que tenía el interior se dice que se ha alcanzado el potencial de acción, que supone la transmisión del impulso nervioso.



La transmisión del impulso nervioso sigue la Ley del todo o nada. Esto quiere decir que si la despolarización de la membrana no alcanza un potencial mínimo, denominado potencial umbral, no se transmite el impulso nervioso, pero, aunque este potencial sea rebasado en mucho, sólo se envía un impulso nervioso, siempre de la misma intensidad.
Una vez que la neurona emite el impulso nervioso debe volver al inicial potencial de reposo. Para ello, la membrana se repolariza, cerrándose los canales para el sodio que estaban abiertos por la presencia del neurotransmisor. El neurotransmisor es destruido por acción enzimática y el potencial de reposo se alcanza al expulsar el sodio la bomba de Na+/K+.
Sinapsis



En las fibras nerviosas amielínicas el impulso se conduce, como una onda continua de inversión de voltaje hasta los botones terminales de los axones. La velocidad que es proporcional al diámetro del axón y varía entre 1 a 100 m/s.
En las fibras nerviosas mielínicas, el axón está cubierto por una vaina de mielina formada por la aposición de una serie de capas de membrana celular, que actúa como un aislante eléctrico del axón.A lo largo del axón, la mielina es formadas por células sucesivas y en cada límite intercelular existe un anillo sin mielina que corresponde al nodo de Ranvier.La consecuencia es una conducción saltatoria del potencial de acción ya que la inversión del voltaje inducido a nivel de un nódulo de Ranvier se continúa por propagación pasiva rápida de la corriente por el interior del axón y por el extracelular hasta el nódulo siguiente donde produce la inversión del voltaje. La consecuencia de esta estructura es que en los axones mielínicos la conducción del impulso nervioso es más rápida. La velocidad de conducción del impulso nervioso es proporcional al diámetro del axón y a la distancia entre los nodos de Ranvier.

Uniones intercelulares de las neuronas
Las neuronas se asocian entre sí y con otras células en estructuras especializadas que permiten:
El tejido nervioso I: tipos de neuronas.
El tejido nervioso II: estructura de la neurona.
El tejido nervioso III: el axón.
El tejido nervioso IV: la mielina en el sistema nervioso periférico.
El tejido nervioso V: la mielina en el sistema nervioso central.
El tejido nervioso VI: estructura de la mielina.
El tejido nervioso VII: la mielinogénesis.
El tejido nervioso VIII: la fibra amielínica.
La sinapsis I: elementos y tipos de sinapsis.
La sinapsis II: histofisiología
La sinapsis III: tipos topográficos y químicos.
La sinapsis IV: la polisinápsis.
La sinapsis V: la sinapsis en barra y bastón.
La sinapsis VI: la sinapsis neuromuscular o placa motora.
Lámina 17- La neurona .
Lámina 18- La membrana y el axon.
Lámina 19- La sinapsis.
Neuronas: (médula, cerebelo, estómago, duodeno)
En el tejido nervioso del SNC, por cada neurona hay entre 10 a 50 células de neuroglia, y que a diferencia de las neuronas retienen su capacidad de proliferar.
Existen 4 clases de células de neuroglia:

 Astrocitos
Astrocitos
Tienen formas estrelladas y presentan largas prolongaciones que se extienden hacia las neuronas y hacia los láminas basales que rodean a los capilares sanguíneos (pies terminales), o que separan al tejido nervioso del conjuntivo laxo de la piamadre, constituyendo la glia limitante
Se han identificado dos tipos de astroglia: astrocitos fibrosos que se asocian de preferencia a las fibras nerviosas de la sustancia blanca y astrocitos protoplasmáticos que se concentran de preferencia asociados a los pericariones, dendritas, terminaciones axónicas en la sustancia gris.
Oligodendrocitos
Son más pequeños y con menos prolongaciones que la astroglía.
Su núcleo es rico en heterocromatina y su citoplasma polirribosomas libres, un aparato de Golgi desarrollado y un alto contenido en microtúbulos, tanto en el citoplasma que rodea al núcleo como en sus prolongaciones. Su función más notable es la formación de la mielina, que rodea a los axones del SNC.
El proceso de mielinización del axón por el oligodendrocito es similar al de la célula de Schwann.
Microglia
Se caracterizan por ser pequeñas, con un denso núcleo alargado y prolongaciones largas y ramificadas.
Células de Schwann
Las células de Schawnn se originan de la cresta neural y acompañan a los axones durante su crecimiento, formando la vaina que cubre a todos los axones del SNP desde su segmento inicial hasta sus terminaciones. Ellas son indispensables para la integridad estructural y funcional del axón.


 Entre las sucesivas células de Schwann existen zonas sin mielina llamadas los nodos de Ranvier. La mielina está compuesta por capas de membrana de la célula de Schwann las cuales se disponen así durante el proceso de mielinización , el cual comienza con la invaginación de un axón, de manera que el axolema se adosa estrechamente a la membrana plasmática de la célula de Schwann por una parte, y las membranas de la célula de Schwann que se enfrentan en el mesaxón. Se produce luego un crecimiento en espiral del citoplasma de la célula de Schwann que se traduce en un crecimiento del mesaxón en forma tal que se enfrentan las membranas plasmáticas de la célula de Schwann por sus caras extracelulares y por sus caras intracelulares. Al fusionare las caras extracelulares se genera la llamada línea interperiodica (línea densa menor) y al desplazarse el citoplasma y fusionarse las caras intracelulares de las membranas se originan las líneas periódicas ( líneas densas mayores)
Entre las sucesivas células de Schwann existen zonas sin mielina llamadas los nodos de Ranvier. La mielina está compuesta por capas de membrana de la célula de Schwann las cuales se disponen así durante el proceso de mielinización , el cual comienza con la invaginación de un axón, de manera que el axolema se adosa estrechamente a la membrana plasmática de la célula de Schwann por una parte, y las membranas de la célula de Schwann que se enfrentan en el mesaxón. Se produce luego un crecimiento en espiral del citoplasma de la célula de Schwann que se traduce en un crecimiento del mesaxón en forma tal que se enfrentan las membranas plasmáticas de la célula de Schwann por sus caras extracelulares y por sus caras intracelulares. Al fusionare las caras extracelulares se genera la llamada línea interperiodica (línea densa menor) y al desplazarse el citoplasma y fusionarse las caras intracelulares de las membranas se originan las líneas periódicas ( líneas densas mayores)
El citoplasma de la célula de Schwann permanece : junto al axón; junto a la superficie externa de la célula y entre las lamelas internodales de la mielina: en l
El astrocito protoplásmico
El astrocito fibroso.
El oligodendrocito.
La microglia y el ependimocito.
Lámina 20- La glía..
Glía: (médula)
Estructura básica del SNP
En el SNP las células nerviosas se organizan formando dos tipos de estructuras:
Los fascículos nerviosos pueden agruparse formando los NERVIOS propiamente tales, en los cuales varios fascículos se agrupan rodeados por el epineuro que es una vaina de tejido conjuntivo que tiende a ser más bien laxo entre los distintos fascículos pero que se hace bastante fibroso en la periferia, estructurando los troncos nerviosos. El epineuro es un tejido conjuntivo corriente, rico en fibroblastos y en fibrillas de colágeno.



ANIMACIONES




impulso nervioso
CUESTIONES: Nervioso
15. SANGRE
 La sangre se presenta como un líquido complejo que sirve para transportar los gases respiratorios y los principales nutrientes celulares, así como para retirar los productos de deshecho. Llena el sistema cardiovascular y se mueve por un camino cerrado o circuito de vasos, gracias a la acción de bombeo del corazón.
La sangre se presenta como un líquido complejo que sirve para transportar los gases respiratorios y los principales nutrientes celulares, así como para retirar los productos de deshecho. Llena el sistema cardiovascular y se mueve por un camino cerrado o circuito de vasos, gracias a la acción de bombeo del corazón.
La sangre es un medio de transporte complejo que realiza servicios vitales:
15.1. Composición de la sangre
La sangre es más que un simple líquido. No sólo está formada por un líquido, sino también por células y fragmentos de células especializadas denominadas plaquetas. la parte líquida de la sangre, es decir, el plasma es uno de los líquidos principales del cuerpo junto con el intersticial y el intracelular. con el término elementos formes designamos los diversos tipos de células de la sangre y los fragmentos celulares suspendidos en el plasma.
El volumen total de sangre varía considerablemente de unos sujetos a otros. la edad, el tipo corporal, el peso, el sexo y el método de medida son fundamentales. para un hombre adulto joven de 70 kg de peso es de unos 5 litros. Una de las principales variables que modifican el volumen de sangre es la cantidad de grasa corporal. el volumen de sangre por kg de peso corporal varía inversamente con el exceso de grasa corporal. ello significa que cuanta menos grasa se tenga en el cuerpo, más sangre habrá por kilogramo de peso corporal.


El plasma es un líquido claro y de color paja. Contiene un90% de agua y un 10% de solutos. La mayor cantidad de solutos son proteínas 6-8% del plasma. Otros solutos, aunque en cantidades menores son: sustancias alimenticias como glucosa y aminoácidos, sustancias formadas en el meta- bolismo como urea, ácido úrico, CO , sustancias reguladoras como hormonas y otros elementos. Las proteínas presentes en el plasmason de tres clases albúminas (55%), globulinas (38%) y fibrinógeno 7%.
Las proteínas plasmáticas son sustancias de importancia decisiva. Por ejemplo, el fibrinógeno y una albúmina desempeñan un papel clave en la coagulación. las globulinas actúan como elementos esen- ciales en los mecanismos de inmunidad; los anticuerpos circulantes son gammaglobulinas modifica- das. Todas las proteínas del plasma contribuyen al mantenimiento de la viscosidad sanguínea nor- mal, de la presión osmótica de la sangre y del volumen sanguíneo. La síntesis de las proteínas del plasma ocurre en el hígado, excepto de las gammaglobulinas, sintetizadas por células plasmáticas.
15.3. Células sanguíneas
Los elementos formes de la sangre son:
Los glóbulos blancos o leucocitos y las plaquetas constituyen menos del 1% del volumen sanguíneo.

 El glóbulo rojo maduro, normal, no tiene núcleo y su diámetro sólo es de unas 7µm, podría colocarse más de 1500, uno al lado del otro, en el espacio de 1 cm. Antes de que la célula alcance la madurez y penetre en el torrente circulatorio desde la médula ósea, el núcleo es extruido, con la consecuencia de que la célula se hunde por las dos caras. Tienen forma de diminutos discos bicóncavos. el eritrocito maduro es también original al no contener ribosomas, mitocondrias ni otros orgánulos típicos de la mayoría de los cuerpos celulares. Por el contrario, el principal componente de cada glóbulo rojo es el pigmento rojo, llamado hemoglobina. Constituye más de la tercera parte del volumen celular.
El glóbulo rojo maduro, normal, no tiene núcleo y su diámetro sólo es de unas 7µm, podría colocarse más de 1500, uno al lado del otro, en el espacio de 1 cm. Antes de que la célula alcance la madurez y penetre en el torrente circulatorio desde la médula ósea, el núcleo es extruido, con la consecuencia de que la célula se hunde por las dos caras. Tienen forma de diminutos discos bicóncavos. el eritrocito maduro es también original al no contener ribosomas, mitocondrias ni otros orgánulos típicos de la mayoría de los cuerpos celulares. Por el contrario, el principal componente de cada glóbulo rojo es el pigmento rojo, llamado hemoglobina. Constituye más de la tercera parte del volumen celular.
La depresión de las superficies planas de la célula da lugar a un centro fino con bordes más gruesos. Esta forma original del hematíe le proporciona una superficie muy grande en relación con su volumen. Los bordes de la célula pueden moverse hacia adentro y hacia afuera permitiendo a esta soportar deformidades o cambios de forma sin que la célula se lesione cuando pase por vasos capilares muy estrechos.
Los hematíes son los más numerosos de los elementos formes en la sangre. En el hombre, su número promedio es de unos 5.500.000 por milímetro cúbico de sangre y en la mujer de 4.800.000 por mm3.
Desempeñan un papel fundamental en el transporte de oxígeno y de dióxido de carbono. Ambas funciones dependen de la hemoglobina.
Encerrados dentro de cada glóbulo rojo hay unos 200- 300 millones de moléculas de hemoglobina, que constituyen aproximadamente el 95% del peso seco de cada célula. cada molécula de hemoglobina está formada, como ya sabéis, por cuatro cadenas de proteína. cada una llamada globina está unida a un pigmento rojo (grupo hemo). cada grupo hemo contiene un átomo de hierro. Por tanto una molécula de hemoglobina contiene cuatro átomos de hierro. Este hecho permite a su vez que cada molécula de hemoglobina se una a cuatro moléculas de oxígeno para formar la oxihemoglobina (reacción reversible).
La hemoglobina puede también unirse al dióxido de carbono formando la carboxihemoglobina (también reversible).
En el eritrocito además de hemoglobina, destaca la presencia de un enzima la anhidrasa carbónica, cataliza la reacción que une el dióxido de carbono con el agua, formando ácido carbónico. la disociación del ácido produce iones bicarbonato (HCO -) que se difunde de los hematíes y actúa transportando CO2. en el plasma sanguíneo. Los iones bicarbonato también desempeñan un importante papel en el mantenimiento de los niveles de pH de la sangre.
La sangre del hombre suele contener más hemoglobina que la de la mujer 14- 16 g/100ml frente a 12- 14. Al adulto que no llega a 10 g/100ml se le diagnostica anemia. las anemias se clasifican de acuedo con el tamaño y el contenido de hemoglobina de los glóbulos rojos.



Formación de los hematíes
El proceso completo en la formación de los glóbulos rojos recibe el nombre de eritropoyesis. En el adulto, los eritrocitos comienzan su secuencia de maduración en la médula ósea roja, partiendo de unas células nucleadas llamadas hemocitoblastos o células madres. las células madre tienen capacidad para mantener una población constante de células en diferenciación. se dividen por mitosis y algunas células hijas siguen como células madre indiferenciadas, mientras que otras atraviesan diferentes fases en el desarrollo hasta transformarse en eritrocitos. El proceso de maduración requiere cuatro días.Los glóbulos rojos se forman y destruyen a una velocidad asombrosa 100 millones por minuto. En estado de salud, el número de glóbulos rojos permanece constante, este número es controlado por mecanismos homeostásicos. El ritmo de formación de hematíes se acelera si disminuye la concentración de oxígeno en sangre que llega a los tejidos. La falta de oxígeno eleva las cifras de glóbulos rojos por aumento de secreción de una hormona glucoproteica denominada eritropoyetina. La eritropoyetina estimula a la médula ósea para que acelere la producción de hematíes. la vida media de los glóbulos rojos circulantes es unos 115 - 120 días Muchas veces a medida que envejecen se fragmentan en los capilares. Los macrófagos se encargan de fagocitar a los
hematíes viejos, anormales o fragmentados. El hierro de la hemoglobina es devuelto a la médula ósea para ser reutilizado en la síntesis de nueva hemoglobina. La bilirrubina es transportada al hígado para su excreción al intestino como parte de la bilis. Los aminoácidos liberados son utiliza- dos por el cuerpo bien para obtener energía o para la síntesis de nuevas proteínas. Para formar nuevos hematíes la médula ósea necesita aportes de vitamina B, hierro, aminoácidos, así como cobre y cobalto que actúan de catalizadores.
GLÓBULOS BLANCOS
 Hay cinco tipos de glóbulos blancos (GB), o leucocitos, clasificados según la presencia o ausencia de gránulos y las características de tinción de su citoplasma.
Hay cinco tipos de glóbulos blancos (GB), o leucocitos, clasificados según la presencia o ausencia de gránulos y las características de tinción de su citoplasma.


Neutrófilos: Con colorantes neutros los gránulos de su citoplasma se tiñen de color púrpura muy claro. Estos gránulos son pequeños y numerosos y tienden a dar al citoplasma un aspecto áspero. Se les denomina también leucocitos polimorfonucleares porque sus núcleos tienen dos, tres o más lóbulos
El número aproximado de neutrófilos es del 65% del total de recuentos de GB en una muestra de sangre normal. Estos leucocitos son sumamente móviles y fagocitos muy activos, que pueden emigrar de los vasos y penetrar en los espacios de los tejidos (DIAPÉDESIS). Los gránulos citoplasmáticos contienen potentes lisosomas, que son orgánulos con enzimas de tipo digestivo capaces de destruir las células bacterianas.
Las infecciones bacterianas que provocan una respuesta inflamatoria causan la liberación de sustancias químicas por las células lesionadas, que atraen neutrófilos y otros GB fagocíticos al lugar de la infección. El proceso, llamado quimiotaxis positiva, ayuda al cuerpo a concentrar células fagocíticas en los focos de infección.
Eosinófilos: Contienen numerosos gránulos citoplasmáticos grandes que se tiñen de naranja con los colorantes ácidos como la eosina. Su núcleo suele tener dos lóbulos. Son el 2 - 5% de los GB circulantes.Son numerosos en algunas células del cuerpo como en el recubrimiento de los paratos respiratorio y digestivo. Aunque son fagocitos débiles, pueden ingerir sustancias químicas y proteínas inflamatorias relacionadas con los complejos antígeno- anticuerpo. Sus funciones más importantes tal vez sean de protección frente a las infecciones causadas por gusanos parásitos y a las reacciones alérgicas.
Basófilos : Son relativamente grandes, pero escasos, y sus gránulos citoplasmáticos se tiñen de rojo púrpura oscuro con los colorante básicos. Son los GB menos numerosos, alcanzando sólo el 0,5-1% del recuento total de leucocitos. Son móviles y capaces de diapédesis. Tienen forma de S, pero sus núcleos son borrosos. Los gránulos citoplasmáticos de estos GB contienen histamina (una sustancia química inflamatoria) y heparina (un anticoagulante).
Linfocitos: Son los leucocitos más pequeños, con un diámetro promedio de 6- 8µm. Poseen núcleos grandes, esféricas, rodeados por una cantidad muy pequeña de citoplasma, que se tiñe azul claro. Son con los neutrófilos los GB más numerosos. Constituyen aproximadamente el 25% de toda la población leucocitaria. Dos tipos de linfocitos, denominados linfocitos T y linfocitos B, desempeñan un papel importante en la inmunidad. Los linfocitos T atacan directamente las células infectadas o cancerosas, mientras que los linfocitos B producen anticuerpos frente a antígenos específicos.
 Monocitos: Son los leucocitos de mayor tamaño. Sus núcleos son oscuros, tienen forma de habichuela y están rodeados de grandes cantidades de citoplasma de color azul-gris característico. Son células móviles y muy fagocíticas, capaces de absorber grandes bacterias y células infectadas por virus.
Monocitos: Son los leucocitos de mayor tamaño. Sus núcleos son oscuros, tienen forma de habichuela y están rodeados de grandes cantidades de citoplasma de color azul-gris característico. Son células móviles y muy fagocíticas, capaces de absorber grandes bacterias y células infectadas por virus.
El número de leucocitos por mm3 es 5000- 9000, con porcentajes distintos de cada clase. Como estas cantidades varían en determinadas situaciones anormales tienen importancia clínica. En la apendicitis aguda, por ejemplo el porcentaje de neutrófilos aumenta, al igual que el recuento total de leucocitos. Denominamos leucopenia a un número excesivamente bajo de GB menos de 5000/ mm3. El SIDA por ejemplo produce una elevada leucopenia. El término leucemia se aplica a un grupo de afecciones malignas caracterizado por la transformación de las células madres que sustituyen a las células normales y provocan leucocitosis (número excesivo de leucocitos) y anemia. Las células leucémicas abandonan la médula ósea y se infiltran por hígado, bazo, SNC, y otras partes del cuerpo.
Formación de leucocitos
La célula madre hemocitoblasto no sólo es precursora de los eritrocitos, sino también de los leucocitos y de las plaquetas de la sangre.
Los granulocitos y algunos linfocitos y monocitos se originan en la médula ósea roja (tejido mieloide). La mayoría de los linfocitos y los monocitos derivan de los hemocitoblastos del tejido linfático. Aunque en la médula ósea se encuentran muchos linfocitos, es probable que la mayoría de ellos se haya formado en tejidos linfáticos y hayan sido transportados a la médula ósea por el torrente circulatorio.
El tejido mieloide y el tejido linfático constituyen los tejidos hematopoyéticos o formadores de células sanguíneas. La médula ósea roja es tejido mieloide en activo, su color rojo proviene de los glóbulos rojos que contiene. La médula ósea amarilla almacena una importante cantidad de grasa, mientras permanece amarilla no es activa en la formación de células sanguíneas, pero puede ser activada cuando se produce una necesidad extrema de hematíes.
PLAQUETAS
Son pequeños cuerpos casi incoloros que por lo general aparecen como discos en forma de huso u ovales que suelen alcanzar 2- 4 µm de diámetro.
Tres propiedades físicas de las plaquetas son: la aglutinación, la adherencia y la agregación. Tan pronto como se extrae sangre de un vaso, las plaquetas se adhieren entre sí y a cualquier superficie con la que entran en contacto; de este modo adoptan formas diversas e irregulares.
El recuento de plaquetas en el adulto arroja un promedio de 250000 por mm3 de sangre. Se consideran normales unos límites de 150.000 – 350000 por mm3. Los niños recién nacidos muestran con frecuencia recuentos más bajos, que aumentan gradualmente para alcanzar los valores normales del adulto hacia los 3 meses de edad. No existen diferencias entre sexos.
 Las plaquetas desarrollan una importante función hemostasia y en la formación de coágulos o coagulación. Aunque relacionadas entre sí, ambas funciones son independientes y distintas.
Las plaquetas desarrollan una importante función hemostasia y en la formación de coágulos o coagulación. Aunque relacionadas entre sí, ambas funciones son independientes y distintas.



Uno a cinco segundos después de la lesión de un capilar, las plaquetas se adhieren al recubrimiento lesionado del vaso y entre sí, formando un tapón hemostático de plaquetas que facilita la detención del paso de sangre a los tejidos. Al menos un tipo de prostaglandinas y ciertas sustancias de origen prostaglandínico, desempeñan un papel en la hemostasia y en la formación de coágulos. Cuando se liberan, estas sustancias actúan sobre el flujo de sangre local (por vasoconstricción) y sobre la agregación plaquetaria en el punto de lesión. Si está es grande, se pone en marcha el mecanismo de formación de coágulos para facilitar la hemostasia.
Las plaquetas se forman en la médula ósea roja, pulmones y, hasta cierto punto, en el bazo por fragmentación de unas células muy grandes (40 – 100 µm), llamadas megacariocitos. estas células se caracterizan por sus grandes núcleos multilobulados que muchas veces tienen forma extraña.
La vida de las plaquetas es corta, unos 9 días por término medio.
15.4. grupos sanguíneos
El término tipo de sangre se refiere al tipo de antígenos (llamados aglutinógenos) presentes en la membrana celular de los hematíes. Los antígenos A, B, y Rh son fundamentales en la sangre en lo referente a las transfusiones y la supervivencia del recién nacido. Se han aislado otros antígenos en la sangre pero su importancia clínica es menor.
SISTEMA ABO
La sangre de todos los seres humanos se corresponde a uno de los cuatro grupos sanguíneos ABO. Estos tipos se denominan de acuerdo con los antígenos presentes en la membrana de los hematíes. He aquí los cuatro tipos ABO:


En la sangre de tipo A el antígeno A está presente en los hematíes, de modo que su plasma no contiene anticuerpos anti- A, pero sí anti- B. En la sangre del grupo B está presente el antígeno B en los hematíes, de manera que el plasma no contiene anticuerpos anti- B, pero sí anti- A. Así pues, la sangre de tipo A donada a un receptor del grupo A no provoca una reacción de aglutinación porque los anticuerpos de tipo B del receptor no se combinan con los antígenos de tipo A de la sangre donada. Sin embargo, la sangre de tipo A donada a un receptor del grupo B provoca una reacción de aglutinación porque los anticuerpos de tipo A del receptor se combinan con los antígenos de tipo A de la sangre donada.
La sangre de tipo O se ha denominado del donante universal, término que significa que se puede administrar con seguridad a cualquier receptor. Sin embargo, esto no es así, ya que el plasma del receptor puede contener aglutininas distintas de los anticuerpo anti-A o anti- B. Por esta razón, la sangre del receptor y del donante cuando son del grupo O deben cruzarse (mezclarse) y observar si existe aglutinación en los hematíes del donante.
La sangre receptora universal (grupo AB) no contiene anticuerpos anti-A ni anti- B, por lo que no puede aglutinar los hematíes de los donantes del tipo a o del tipo B. Ello no significa que cualquier tipo de sangre donada pueda administrarse con seguridad a un individuo con sangre del grupo AB sin cruzarla previamente.
En la sangre del llamado receptor universal, puede haber otras aglutininas que aglutinen antígenos no identificados (aglutinógenos) de la sangre del donante.
SISTEMA Rh
El término Rh positiva significa que está presente el antígeno Rh en sus hematíes. Por otra parte, en la sangre Rh negativa los hematíes no tienen antígeno Rh.
Por lo general ninguna sangre contiene anticuerpos Rh, aunque pueden aparecer en la sangre de las personas Rh negativo si en alguna ocasión han entrado hematíes Rh positivos en el torrente circulatorio. Una forma en que esto puede pasar es administrar una transfusión de Rh + a una persona Rh-. Al cabo de poco tiempo esa persona produce anticuerpos que quedan en la sangre. La otra vía por la que hematíes Rh+ pueden entrar en el torrente circulatorio de un Rh- es durante el embarazo. En ello reside el riesgo de un bebé nacido de una madre Rh- y del padre Rh+. Si el niño hereda ser Rh+ del padre, el factor Rh de los hematíes puede estimular el cuerpo de la madre para que forme anticuerpos anti- R. Por lo tanto si más tarde queda embarazada de otro feto Rh+, este puede padecer una enfermedad llamada eritrobastosis fetal, producida por la reacción entre los anticuerpos Rh de la madre y las células Rh+ del feto. Algunas madres Rh- embarazadas de niños Rh+ son tratadas con una proteína que impide que el cuerpo de la madre forme anticuerpos anti- Rh, evitando así la posibilidad de lesión al siguiente niño Rh+. (eritroblastosis fetal)
Los cromosomas y los genes se localizan por parejas, ya que proceden uno del padre y el otro de la madre. Así, los diferentes genotipos para los distintos grupos sanguíneos son :
-IA IA es la dotación génica responsable del grupo A;
-IA i es la dotación génica híbrida del grupo A;
-IB IB es la dotación génica responsable del grupo B;
-IB i es la dotación génica híbrida del grupo B;
-IA IB es la dotación génica responsable del grupo AB;
-i i es la dotación génica responsable del grupo O.
Lámina 9 .- Celulas inmunocompetentes.
Lámina 10.- Células de la sangre.
Elementos figurados
Las células de la sangre I: los granulocitos.
Las células de la sangre II: el eritrocito
Las células de la sangre III: los agranulocitos.
Las células de la sangre IV: la granulocitopoyesis .
Las células de la sangre V: eritropoyesis.
Las células de la sangre V: plaquetopoyesis.
16. RESUMEN
Tejidos animales
PORTADA
7.1.1. EPITELIO PLANO MONOESTRATIFICADO
Las células de los epitelios planos monoestratificados son aplanadas y tienen bordes sinuosos mutuamente interdigitados. Frecuentemente, la región celular en donde se aloja el núcleo está abombada por su presencia.Se encuentra tapizando parte de las nefronas (que estudiaremos en el capítulo correspondiente al aparato excretor), recubriendo la cámara anterior del ojo, la pared de los alvéolos pulmonares y la cara interna del tímpano.
Existen dos tipos especiales de epitelios planos monoestratificados:
- Endotelio: se designa al epitelio plano monoestratificado que tapiza la cara interna de los vasos sanguíneos y linfáticos y las cavidades del corazón (aurículas y ventrículos).
- Mesotelio: es el que forma parte de las membranas serosas que recubren las tres grandes cavidades del cuerpo:
- Peritoneal: en donde se aloja el aparato digestivo, urogenital, ...
- Torácica: en donde se encuentra el aparato respiratorio.
- Pericárdica: que contiene el corazón
7.1.2. EPITELIO CÚBICO SIMPLE
Formado por una simple capa de células c†úbicas apoyadas en la lá€mina basal. Se encuentra
tapizando los conductos que vierten el contenido de algunas glándulas al exterior.
7.1.3. EPITELIO CILÍNDRICO MONOESTRATIFICADO
Está muy extendido en el cuerpo humano. Se encuentra tapizando zonas de nuestro cuerpo con actividad secretora y de absorción, como el caso del aparato digestivo.Algunos epitelios de este tipo presentan cilios en la zona apical, como el epitelio que recubre el interior de la vesícula biliar o los oviductos.
Otros epitelios cilíndricos monoestratificados contienen otros tipos celulares, como la células caliciformes, glándulas unicelulares que fabrican secreciones mucosas y que estudiaremos más adelante.
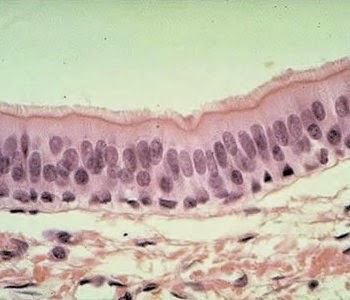
7.1.4. EPITELIO SEUDOESTRATIFICADO CILÍNDRICO
Como ya sabemos, estos epitelios est€án formados por células redondeadas y en forma discontinua que no llegan todas a la superficie, y cé‚lulas prismá€ticas de 40 a 50 cm. de altura. Suelen tener cilios. Se localizan en la mucosa pituitaria (fosa nasal), bronquios, algunas regiones de la uretra masculina y vías excretoras del aparato genital masculino.
7.1.5. EPITELIO PLANO PLURIESTRATIFICADO
En las zonas sometidas a un esfuerzo mecánico intenso, se encuentra el epitelio plano estratificado como tejido protector. En las superficies interiores, cavidad bucal, vagina y ano, no est€á cornificado.
El epitelio plano estratificado cornificado forma la epidermis o estrato superior de la piel. Protege al organismo contra agentes nocivos físicos y quíƒmicos, e impide la desecación de la piel.
En la epidermis de la piel se distinguen cinco estratos: el estrato basal, el estrato espinoso, el estrato granuloso, el estrato lúcido y el estrato córneo.
El estrato basal, que también se llama estrato germinativo porque contiene células en división, es adyacente a la lámina basal. Las cé‚lulas nuevas producto de mitosis van siendo desplazadas hacia la superficie para reemplazar a aquellas que se han descamado.
El estrato espinoso tiene varias células de espesor. Las células de esta capa poseen “espinas” en su superficie que están en contacto con formaciones similares en células vecinas. Al microscopio electrónico se observa que los puntos de contacto entre dos espinas de dos c‚élulas adyacentes son, en realidad, desmosomas.
En las zonas sometidas a un esfuerzo mecánico intenso, se encuentra el epitelio plano estratificado como tejido protector. En las superficies interiores, cavidad bucal, vagina y ano, no est€á cornificado.
El epitelio plano estratificado cornificado forma la epidermis o estrato superior de la piel. Protege al organismo contra agentes nocivos físicos y quíƒmicos, e impide la desecación de la piel.
En la epidermis de la piel se distinguen cinco estratos: el estrato basal, el estrato espinoso, el estrato granuloso, el estrato lúcido y el estrato córneo.
El estrato basal, que también se llama estrato germinativo porque contiene células en división, es adyacente a la lámina basal. Las cé‚lulas nuevas producto de mitosis van siendo desplazadas hacia la superficie para reemplazar a aquellas que se han descamado.
El estrato espinoso tiene varias células de espesor. Las células de esta capa poseen “espinas” en su superficie que están en contacto con formaciones similares en células vecinas. Al microscopio electrónico se observa que los puntos de contacto entre dos espinas de dos c‚élulas adyacentes son, en realidad, desmosomas.
La producción continua de nuevas células en el estrato basal hace que las cé‚lulas del estrato espinoso suprayacente sean empujadas hacia el estrato granuloso. Durante este movimiento las c‚élulas se aplanan quedando paralelas a la superficie de la piel.
El estrato granuloso se distingue debido a que sus c‚élulas contienen numerosos gránulos de queratina, los cuales se tiñen como hematoxilina.
El estrato córneo es la capa superficial. Las cé‚lulas en este estrato ya no poseen ni n†cleo ni orgánulos. La membrana plasm€ática estဠengrosada y cubierta en su superficie externa por glucolƒípidos; el interior de las células esta completamente lleno de queratina.
7.1.6. EPITELIO DE TRANSICIÓN
Representa una forma especial de epitelio cilíndrico estratificado que solamente existe en las vías urinarias. Este epitelio puede modificar su forma por deslizamiento de unas cé‚lulas sobre todas. Cuando la vƒíscera a la que pertenecen está distendida, el epitelio sólo consta de hasta diez o más capas de células cúbicas y de otras formas. A medida que aumenta la tensión, la capa epitelial se extiende, el número de capas celulares visibles disminuye y la forma de las célula pasará de cúbica a planas. Este epitelio se encuentra en la vejiga urinaria y la protege de posibles desgarros. FUNCIONES DE LOS EPITELIOS DE REVESTIMIENTO
- Protección: Todos los tipos de epitelios de revestimiento, en principio, ejercen una función de protección; es decir, no sólo el epitelio estratificado plano queratinizado est€á especializado en ese sentido aunque sea paradigm€tico. Las escamas de queratina le confieren la capacidad de resistencia mecá€nica y quƒímica e impiden la desecación del organismo. Los grá€nulos de melanina procedentes de los melanocitos se encuentran distribuidos por la epidermis y son capaces de absorber los rayos ultravioleta, por lo que protegen al organismo del efecto perjudicial de las radiaciones.
- Absorción: Es la principal tarea del epitelio simple cilƒndrico del intestino delgado. En la figura se aprecia la absorción del producto de la digestión de grasas que son de nuevo resintetizados en el interior de la c‚élula epitelial y combinadas con proteƒnas para formar quilomicrones solubles que son lanzados al vaso linf€ático quilífero
- Movimiento: Conseguido mediante cilios.
- Actividad secretora: En forma de c‚élulas especializadas intercaladas entre las normales de revestimiento.
- Deslizamiento: Los mesotelios, tienen como tarea usual el asegurar el deslizamiento entre los órganos internos.
- Intercambio gaseoso: El epitelio simple plano de los alvéolos pulmonares interviene en el intercambio gaseoso entre la sangre y la luz alveolar. El dióxido de carbono de la sangre se elimina a trav‚és de la pared capilar y de la delgada zona citoplasm€ática de las c‚élulas alveolares, mientras que el oxígeno se difunde a la sangre por el mismo camino.
Resumen de tejidos epiteliales de revestimiento
|
|||
Tipos
|
Características
|
Función
|
Ejemplos
|
Planos
|
Células muy delgadas
|
Intercambio
|
Pulmones
Vasos sanguíneos |
Cúbicos
|
Células prismáticas
|
Conducción
|
Colector renal
|
Cilíndricos
|
Células prismáticas alargadas
|
Absorción. Protección
|
Intestino
Epidermis muchos invertebrados |
Pluriestratificado plano
|
Varias capas planas
|
Protección
|
Piel
|
Pluriestratificado cúbico
|
Varias capas cúbicas
|
ConducciónContención
|
Conductos sudoríaros
Vegiga |
Pluriestratificado cilíndrico
|
Varias capas cilíndricas
|
Conducción
|
Conductos mamarias
|
Pseudoestratificado
|
Como en dos estratos no nítidos
|
Conducción
|
Tráquea
|
Epitelial. De revestimiento. Simple
- Plano: (esófago, lengua)
- De transición: (vejiga urinaria)
- Seudoestratificado: (tráquea)
- Células aisladas: (duodeno)
- Tubular: (estómago)
- Acinar: (glándula salivar, páncreas, tráquea)
Las glándulas son asociaciones de células epiteliales altamente diferenciadas, que segregan sus productos de síntesis, bien a una superficie libre, mediante un conducto secretor (glándulas exocrinas), o bien directamente al torrente sanguíneo (glándulas endocrinas o de secreción interna).
Todas las células del organismo del organismo están dotadas de actividad secretora. La pro- piedad de secreción sólo se convierte en función cuando moviliza total o casi totalmente la actividad de la célula y cuando conduce a la elaboración de productos utilizados por el orga- nismo. Una célula capacitada para tal función tiene una actividad glandular
La secreción se desarrolla según un proceso constante cualquiera que sea el tipo celular. Comprende una sucesión de cuatro fases:
- Una fase de asimilación de las unidades necesarias para la síntesis; la célula toma líquido del medio intersticial .
- Una fase de síntesis o elaboración del producto de secreción, durante la cual la célula asocia los precursores en un orden genéticamente predeterminado.
- Una fase de “embalaje” y almacenamiento (no constante) del producto de secreción, durante la cual aumenta la concentración de éste.
- Fase de secreción o descarga, durante la cual el producto elaborado es expulsado de la célula.
Las glándulas se forman a partir de superficies epiteliales. El epitelio prolifera y se hunde, en el tejido conjuntivo, manteniendo, o no, contacto con la superficie, según que vaya a ser glándula exocrina o endocrina, respectivamente. En las glándulas endocrinas, las células pueden agruparse en cordones o en folículos. En los folículos, el producto de secreción se acumula en gran cantidad. En las Glándulas exocrinas el producto se vierte al medio externo bien por un conducto excretor, bien directamente. El medio externo puede ser:
- La superficie cutánea (glándulas sebáceas, sudoríparas, etc.)
- La luz de una cavidad natural (glándulas digestivas, genitourinarias, del aparato respi- ratorio...).
- Directamente a partir de células glandulares (hipófisis, glándulas suprarrenales)
- Indirectamente después de un periodo de almacenamiento extracelular (tiroides)
Según la forma de secreción las glándulas se clasifican en:
Merocrinas: que es una forma de secreción que respeta la integridad de la células, es decir, ésta no muere ni sale dañada en el proceso. la mayoría de las glándulas pertenecen a este tipo (salivares, páncreas, etc.)
Apocrinas: es una forma de secreción en la cual el producto de secreción acumulado en el polo apical de la célula se expulsa con éste. Las células así decapitadas regeneran rápidamente esta pérdida de
sustancia y comienzan nuevamente un ciclo secretor. Son ejemplos típicos de glándulas apocrinas, las glándulas mamarias, algunas glándulas su- doríparas del hueco axilar, de los pliegues inguinales, de los órganos genitales, de la región perianal y de los párpados (las llamadas glándulas de Moll)
Holocrinas: tipo de secreción en la que la cesión de sustancias lleva consigo la muerte celular. En este caso, la célula está completamente llena de se- creción. La secreción holocrina tiene lugar, sobre todo, en las glándulas sebáceas de la piel. Las glándulas también pueden clasificarse atendiendo al número de células y disposición de
Glándulas unicelulares: la glándula es una sola célula. Como ejemplo cita- remos a las ya mencionadas glándulas o células caliciformes. Estas células fabricantes de moco y glucoproteínas tienen forma de copa y se alternan entre las células de un epitelio de revestimiento. Vierten su contenido al exterior para formar una capa protectora.
Glándulas pluricelulares: se dividen en simples, ramificadas y compuestas. En las glándulas ramificadas, varias unidades secretoras individuales desembocan en un conducto excretor común. En las glándulas compuestas, el conducto secretor principal está subdividido en varios conductos más pequeños.
Según el aspecto de la unidades glandulares secretoras, también llamadas unidades glandulares , las glándulas se clasifican también como acinosas (en forma de acino, como frutos en baya), alveolares (en forma de sáculos) y tubulares (en forma alargada, como un tubo). Existen formas mixtas que son glándulas tubuloacinosas y las tubuloalveolares.
Las unidades glandulares están formadas por células secretoras rodeadas por una lámina basal.
Los epitelios sensoriales, altamente especializados, intervienen en la captación de impresiones sensoriales, serán tratados en el tema dedicado a los sentidos.
8. TEJIDOS CONECTIVOS
Los tejidos conjuntivos , derivados del mesénquima, constituyen una familia de tejidos que se caracterizan porque sus células están inmersas en un abundante material intercelular, llamado la matriz extracelular.
Existen 2 variedades de células conjuntivas:
Existen 2 variedades de células conjuntivas:
- Células estables, las que se originan en el mismo tejido y que sintetizan los diversos componentes de la matriz extracelular que las rodea
- Una población de células migratorias, originadas en otros territorios del organismo, las que llegan a habitar transitoriamente el tejido conjuntivo.
- Mantener unidos entre sí a los otros tejidos del individuo.
- Contener a las células que participan en los procesos de defensa ante agente extraños: constituyendo el sitio donde se inicia la reacción inflamatoria.
- Constituir un medio tisular adecuado para alojar células en proceso de proliferación y diferenciación para formar los elementos de la sangre correspondientes a glóbulos rojos y plaquetas, y a los distintos tipos de glóbulos blancos, los que migran luego a los tejidos conjuntivos, para realizar en ellos sus funciones específicas ya sea como células cebadas, macrófagos, células plasmáticas, linfocitos y granulocitos
- Almacenar grasas, para su uso posterior como fuente de energía, ya sea por ellos mismos o para otros tejidos del organismo: Tejido adiposo.
- formar láminas con una gran resistencia a la tracción, tal como ocurre en la dermis de la piel, y en los tendones y ligamentos: Tejido conjuntivo fibroso denso.
- formar placas o láminas relativamente sólidas, caracterizadas por una gran resistencia a la compresión: Tejido cartilaginoso.
- formar el principal tejido de soporte del organismo, caracterizado por su gran resistencia tanto a la tracción como a la compresión: Tejido óseo.
Los tejidos conectivos se pueden clasificar en los siguientes grupos:

 Como ya se ha dicho, los tejidos conectivos tienen tres elementos básicos:
Como ya se ha dicho, los tejidos conectivos tienen tres elementos básicos:
Las células del tejido conectivo están más o menos distanciadas entre sí. La estructura de este tejido se adapta, en cada caso, a las características o exigencias mecánicas del órgano en cuestión. Los tejidos conectivos se encuentran distribuidos por la totalidad del organismo:


LA SUSTANCIA FUNDAMENTAL
La sustancia fundamental es homogénea y amorfa, en estado de gel, y ocupa los espacios comprendidos entre las fibras y las células. Al microscopio electrónico tiene poca densidad (aparece clara)
Contiene principalmente:
FIBRAS DEL TEJIDO CONECTIVO
Las fibras del tejido conectivo determinan el tipo de tejido conjuntivo según su propor- ción y disposición. Aunque hoy las fibras de reticulina se consideran como una variación de las
de colágeno, las estudiaremos por separado. Todas ellas son de naturaleza proteica.
Fibras de colágeno
Etimológicamente, las fibras denominadas “colágenas” son las que, tratadas con agua caliente, dan origen a la gelatina (colas). Estas fibras están formadas por una proteína, el colágeno (ya estudiada), que representa entre un 30 y un 35 % de las proteínas totales del organismo humano.
Las fibras de colágeno son elementos en forma de cinta o de cilindro, con un diámetro que varía de 1 a 5 m y pueden formar haces de hasta más de 13 m. Confieren al tejido conectivo su resistencia a las fuerzas mecánicas y su solidez. laas propiedades de las fibras de colágeno son:



- Tejido conjuntivo propiamente dicho:
- Tejido conjuntivo fibroso laxo.
- Tejido conjuntivo fibroso denso.
- Tejido conjuntivo especializado:
- Tejido conjuntivo denso alineado (ligamientos y tendones).
- Tejido adiposo.
- Tejido sanguíneo. (se verá en el tema de Hematología)
- Tejido óseo.
- Tejido cartilaginoso.
- Tejido hematopoyético.
- Tejido linfoide.
- Tejido conjuntivo embrionario:
- Mesénquima.
- Tejido conectivo mucoso.
- Sustancia fundamental, líquida y amorfa.
- Fibras, que se clasifican en:
- Fibras de colágeno.
- Fibras de reticulina.
- Fibras elásticas.
- Células del tejido conjuntivo.
Las células del tejido conectivo están más o menos distanciadas entre sí. La estructura de este tejido se adapta, en cada caso, a las características o exigencias mecánicas del órgano en cuestión. Los tejidos conectivos se encuentran distribuidos por la totalidad del organismo:
- Rodean órganos, dándoles una naturaleza más consistente.
- Llenan espacios comprendidos entre órganos.
- Constituyen el estroma (sustancia o matriz fundamental de un órgano y no tiene nada que ver con el estroma de un cloroplasto) de un gran número de órganos. Participan en la composición de diferentes aparatos (por ejemplo, aparato circulatorio aparato locomotor,...)
La sustancia fundamental es homogénea y amorfa, en estado de gel, y ocupa los espacios comprendidos entre las fibras y las células. Al microscopio electrónico tiene poca densidad (aparece clara)
Contiene principalmente:
- Sustancias sintetizadas por las células conectivas.
- Como los proteoglicanos o glucosaminoglucanos, que son cadenas polipeptídi- cas que tienen disacáridos aminados como:
- Acido hilurónico. (las bacterias que produce hialuronidasa se despla- zan bien por el tejido conjuntivo)
- Condroitina.
- También glucoproteínas estructurales.
- Sustancias de origen exógeno (procedentes del plasma sanguíneo) como agua, cationes y algunas proteínas como albúminas e inmunoglobulinas.
FIBRAS DEL TEJIDO CONECTIVO
Las fibras del tejido conectivo determinan el tipo de tejido conjuntivo según su propor- ción y disposición. Aunque hoy las fibras de reticulina se consideran como una variación de las
de colágeno, las estudiaremos por separado. Todas ellas son de naturaleza proteica.
Fibras de colágeno
Etimológicamente, las fibras denominadas “colágenas” son las que, tratadas con agua caliente, dan origen a la gelatina (colas). Estas fibras están formadas por una proteína, el colágeno (ya estudiada), que representa entre un 30 y un 35 % de las proteínas totales del organismo humano.
Las fibras de colágeno son elementos en forma de cinta o de cilindro, con un diámetro que varía de 1 a 5 m y pueden formar haces de hasta más de 13 m. Confieren al tejido conectivo su resistencia a las fuerzas mecánicas y su solidez. laas propiedades de las fibras de colágeno son: - Resistentes: a las tracciones y a las fuerzas mecánicas (de ahí su abundancia en cápsulas articulares, tendones y ligamientos). Pueden soportar varios cientos de kg/cm2)
- Flexibles: lo que implica su presencia en la piel (dermis)
- Insolubles en agua fría pero solubles en agua caliente (gelatina)
- Digeribles: por enzimas proteolíticos como proteasas y colagenasas que tenemos en el tracto digestivo.
Fibras de reticulina
Son fibras de colágeno pero mucho más delgadas y se disponen formando redes. A partir de ellas se pueden formar fibras de colágeno. Se encuentran en:
Tienen estructura filamentosa ramificada y forman redes tridimensionales. En contraste con las fibras de colágeno inextensibles, son muy deformables elásticamente. La elastina es la proteína más importante de las fibras elásticas. Se encuentran en casi todos los tejidos conectivos. Las arterias próximas al corazón y a los pulmones, presentan gran abundancia de estas fibras, ya que deben soportar la gran presión con que la sangre sale del corazón. También son abundan- tes en el tejido cutáneo.
Células conjuntivas
Las células que están presente, en en los tejidos conjuntivos corresponden a dos grupos distintos:
Son fibras de colágeno pero mucho más delgadas y se disponen formando redes. A partir de ellas se pueden formar fibras de colágeno. Se encuentran en:
- Tejido conjuntivo embrionario, y en todas las regiones del organismo donde se realiza la síntesis de colágeno.
- En órganos hematopoyéticos: (=órganos relacionados con la producción de células sanguíneas), como el bazo, ganglios linfáticos y médula ósea.
- Alrededor de células glandulares, como las que hay en el riñón, hígado y en las glándulas endocrinas.
- Formando parte de las láminas basales.
Tienen estructura filamentosa ramificada y forman redes tridimensionales. En contraste con las fibras de colágeno inextensibles, son muy deformables elásticamente. La elastina es la proteína más importante de las fibras elásticas. Se encuentran en casi todos los tejidos conectivos. Las arterias próximas al corazón y a los pulmones, presentan gran abundancia de estas fibras, ya que deben soportar la gran presión con que la sangre sale del corazón. También son abundan- tes en el tejido cutáneo.
Células conjuntivas
Las células que están presente, en en los tejidos conjuntivos corresponden a dos grupos distintos:
- Células propias de cada tipo de tejido conjuntivo, las que constituyen una población celular estable, responsable de la formación y mantenimiento del tejido al que pertenecen, razón por la cuál se las llama células de sostén .
- Células conjuntivas libres, que han llegado a residir en los tejidos conjuntivos. Estas células se forman en los órganos hematopoyéticos, circulan como glóbulos blancos en la sangre, y migran luego a un tejido conjuntivo en particular, estableciéndose en él.
- Células de vida media larga, capaces de dividirse en el tejido conjuntivo que habitan: células cebadas y fagocitos mononucleares o macrófagos
- Células de vida media corta, que no se dividen, y cuya concentración en una a zona del tejido conjuntivo se relaciona con procesos de defensa del organismos: células plasmáticas, linfocitos y fagocitos porlimorfonucleares .
Células mesenquimatosas: Son células procedentes del mesodermo presentes en el mesénquima o tejido conjuntivo embrionario. Se puede decir que constituyen un tipo de fibroblasto primitivo o su precursor. Apare- cen también en el tejido mucoide del cordón umbilical. Estas células pueden diferenciarse hacia fibroblastos como ya hemos dicho, adipocitos, células reticulares, miofibroblastos, células musculares lisas o hasta células de Leydig.
Células reticulares: Únicamente se hallan presentes en el tejido conjuntivo reticular que forma parte del estroma de los órganos linfoides y hematopoyéticos.
Reciben este nombre por sus prolongaciones estrelladas por las que se unen a otras células
gracias a desmosomas.
Fibroblastos: Son las células características del tejido conjuntivo y están presentes en la mayor parte de sus variedades y, sobre todo, en las que se encuentran en pleno desarrollo. Se distribuyen en el tejido conjuntivo entre las fibras de colágeno que ellos mismos fabrican. También fabrican la sustancia fundamental.
En general, los fibroblastos tienen forma irregu- lar: son células aplanadas y fusiformes, provis- tas de prolongaciones delgadas que entran temporalmente en contacto con las de los fibro- blastos contiguos. Existen dos tipos de fibroblas- tos y todas las fórmulas intermedias:
El fibroblasto típico o fibroblasto jo- ven, con una intensa actividad de síntesis. Éste tiene capacidad de dividirse si es conveniente- mente estimulado, por ejemplo cuando hay ne- cesidad de cicatrización.
El fibroblasto maduro o fibrocito, es poco activo. El fibroblasto tiene un núcleo de gran tamaño, ovoide y con un nucleolo voluminoso. El citoplasma contiene:
- Retículo endoplasmático en forma de gránulos y no sáculos.
- Aparato de Golgi muy desarrollado.
- Numerosas mitocondrias.
- Lisosomas.
- Abundantes microtúbulos y microfilamentos que le confieren esa forma ramificada.
- Los histiocitos o macrófagos que se encuentran en el tejido conjuntivo.
- Las células de Kupffer del hígado.
- Células de la glía, microglía, del sistema nervioso central.
- Macrófagos de la médula ósea.
- Células de la glía, microglía, del sistema nervioso central.
- Una función de fagocitosis muy desarrollada, bacterias, virus, restos de células muertas, células extrañas,...
- Presenta el antígeno (cualquier célula o sustancia que nuestro organismo reconoce como extraña), a las células que fabricarán los anticuerpos necesarios (linfocitos). Si el antígeno entra en contacto con el linfocito, este no sabrá que anticuerpo fabricar si no es presentado por un macrófago.
Células cebadas o mastocitos: Estas células son muy parecidas a los leucocitos basófilos y provienen de células de la médula ósea. Suelen encontrarse junto a los vasos sanguíneos de pequeño calibre. Son redondeadas, ovoides o fusiformes. Su principal característica es la presencia en el citoplasma de abundante gránulos de unas 0’6 m de diámetro rodeados por membrana.
Estas células intervienen en reacciones de anafilaxia en relación con las inmunoglobulinas de IgE de modo que cuando éstas se unen a las membranas de los masto- citos liberan al exterior el contenido de sus gránulos:
Intervienen también en el reconocimiento celular, crecimiento y regeneración del tejido conjun- tivo, en la respuesta inflamatoria y en el rechazo agudo de trasplantes.
Estas células intervienen en reacciones de anafilaxia en relación con las inmunoglobulinas de IgE de modo que cuando éstas se unen a las membranas de los masto- citos liberan al exterior el contenido de sus gránulos:
- Heparina: un mucopolisacárido anticoagulante.
- Histamina: aumenta la permeabilidad capilar y la presión arterial.
- Serotonina: vasoconstrictor que aumenta la presión arterial por constricción del músculoliso. (esta última sustancia no se ha encontrado en células cebadas del hombre pero sí de roedores).
- Factor quimiotáctico para los leucocitos eosinófilos.
Intervienen también en el reconocimiento celular, crecimiento y regeneración del tejido conjun- tivo, en la respuesta inflamatoria y en el rechazo agudo de trasplantes.
- Monocitos que se transformarán en macrófagos.
- Linfocitos especialmente células plasmáticas.
- Leucocitos neutrófilos polimorfonucleares.
- Leucocitos eosinófilos.
Unas células particulares, Los adipocitos: Son células conjuntivas que acumulan, en forma de reserva, importantes cantidades de lípidos. Son las células típicas del TEJIDO ADIPOSO y existen dos tipos: - Adipocitos de grasa blanca.
- Adipocitos de grasa parda, que en el ser humano sólo existen durante la vida fetal.
- El adipocito es una célula fija, ovoide o esférica, de un diámetro variable entre 50-150 m. Todo su citoplasma está prácticamente ocupado por una vacuola de un con- tenido semilíquido a 37ºC y constituido por una mezcla de:
- Acilglicéridos y ácidos grasos.
- Acidos grasos libre.
- Carotenoides.
Los adipocitos en el tejido conjuntivo cumplen:
- Una función nutritiva.
- Una función de protección mecánica.
- Una función de aislamiento térmico.
El tejido conectivo I: La circulación del agua.
El tejido conectivo II: Las fibras.
El tejido conectivo III: Las células.
El tejido conectivo IV: La síntesis de fibras colágenas.
Lámina 6. Fibras del tejido conectivo.
Lámina 7.- Células del tejido conectivo.
Conectivo típico
Laxo: (esófago, estómago, tráquea, arteria, vena)
Regular: (tendón)
Estudiemos a continuación los distintos tipos de tejido conjuntivo típico
9.1. MESÉNQUIMA
Se denomina mesénquima al tejido conjuntivo embrionario que existe durante el desarrollo intrauterino. Descendientes del mesénquima son, además de las células del tejido conjuntivo, la células musculares lisas. Las células mesenquimatosas se caracterizan por su animada actividad mitótica y porque no forman fibras.
 El tejido gelatinoso está emparentado y es parecido al mesénquima y, asimismo, su existencia es transitoria. Es el tejido típico del cordón umbilical.
El tejido gelatinoso está emparentado y es parecido al mesénquima y, asimismo, su existencia es transitoria. Es el tejido típico del cordón umbilical.
9.2. TEJIDO CONJUNTIVO FIBROSO LAXO O AREOLAR
Es muy frecuente en el organismo y está caracterizado por tener poca densidad de fibras. Se puede decir que se trata de un tejido de relleno.
Forma parte del tejido subcutáneo, sostiene los tejidos epiteliales (por debajo de la lámina basal), constituye el estroma (sustancia fundamental) de numerosos órganos, el iris y coroides del ojo (tejido conjuntivo fibroso laxo pigmentario) y envolviendo haces de músculos y nervios (tejido conjuntivo fibroso laxo laminar). También se encuentra bajo el mesotelio de las serosas (pleura, peritoneo y pericardio)
Este tejido presenta todos los componentes típicos del tejido conectivo, pero las fibras de colágeno (las más numerosas) elásticas y de reticulina sin ninguna orientación en particular.
9.1. MESÉNQUIMA
Se denomina mesénquima al tejido conjuntivo embrionario que existe durante el desarrollo intrauterino. Descendientes del mesénquima son, además de las células del tejido conjuntivo, la células musculares lisas. Las células mesenquimatosas se caracterizan por su animada actividad mitótica y porque no forman fibras.
El tejido gelatinoso está emparentado y es parecido al mesénquima y, asimismo, su existencia es transitoria. Es el tejido típico del cordón umbilical.
Tipos
|
Características
|
Función
|
Localización
|
Laxo
|
Pocas fibras.
|
Relleno. Movimiento
|
Haces musculares. Dermis
|
Denso
|
Gran cantidad de fibras colágenas
|
Resistencia a tracción
|
Tendones. Ligamentos. Dermis
|
Adiposo
|
Muchos adipocitos
|
Reserva. Aislamiento
|
Dermis. Cojinetes. Tras riñones
|
9.2. TEJIDO CONJUNTIVO FIBROSO LAXO O AREOLAR
Es muy frecuente en el organismo y está caracterizado por tener poca densidad de fibras. Se puede decir que se trata de un tejido de relleno.
Forma parte del tejido subcutáneo, sostiene los tejidos epiteliales (por debajo de la lámina basal), constituye el estroma (sustancia fundamental) de numerosos órganos, el iris y coroides del ojo (tejido conjuntivo fibroso laxo pigmentario) y envolviendo haces de músculos y nervios (tejido conjuntivo fibroso laxo laminar). También se encuentra bajo el mesotelio de las serosas (pleura, peritoneo y pericardio)
Este tejido presenta todos los componentes típicos del tejido conectivo, pero las fibras de colágeno (las más numerosas) elásticas y de reticulina sin ninguna orientación en particular.
Las células más nmerosas son los fibtoblastos y los macrófagos.
Las funciones de este tejido son:
- Función mecánica: desempeña el importante papel, al ser deformable y más o me- nos elástico, la movilidad de los tejidos y órganos pero asegurando el regreso de las estructuras a su posición inicial.
- Función de cicatrización: tras la agresión debido a bacterias o lesiones, los fibroblas- tos de este tejido regeneran las pérdidas de sustancia.
- Transporte de metabolitos: permite el paso de nutrientes, oxígeno, anhídrido carbó-
- nico y residuos entre los capilares y las células. Al tener poca sustancia intercelular no hay obstáculos para estos intercambios.
- Función de defensa: corre a cargo de los macrófagos exclusivamente.
9.3. TEJIDO CONJUNTIVO FIBROSO DENSO
Este tipo de tejido se caracteriza por la abundancia de fibras de colágeno y de elastina, y por la escasez de células. Los fascículos de fibras de colágeno siguen trayectos que dependen de la dirección de las fuerzas que tiene que soportar el órgano en cuestión. Se encuentran dos tipos de tejido:
A) TEJIDO CONJUNTIVO FIBROSO DENSO NO ALINEADO
En él, las fibras de colágeno no están orientadas y se distribuyen de manera compacta. Estos tejidos están sometidos a fuerzas multidireccionales. Se encuentran en la duramadre (una meninge = membrana que hay entre el cráneo y el encéfalo), en las articulaciones, en la esclerótica del ojo, formando parte de la cápsula (parte más externa de un órgano y que le da la forma) del testículo, bazo, riñón, hígado y otros. También forma la vaina de los tendones ( un tendón es un cordón fibroso de color blanco nacarado mediante el cuál el músculo se une al hueso o a otro órgano) y el periostio.
Sus fibras de colágeno se encuentran todas alineadas en el sentido de la tracción. Los fibroblastos de este tejido reciben el nombre de células tendinosas. Este tejido está adaptado para transmitir fuerzas de tracción mecánica: en efecto, es poco extensible (5%) y muy resistente (100 kg por cm2 de sección). Se localiza en los tendones (que transmiten a los
huesos el efecto de tracción desarrollados por los músculos) y en los ligamentos (un ligamento es una cinta o fascículo que se inserta en huesos y cartílagos y que sirve para como medio de unión de la articulaciones o para otros fines). Es también un tejido típico de la córnea.
9.4. TEJIDO CONJUNTIVO ELÁSTICO
Es un tejido rico en fibras elásticas aunque también hay de colágeno. Las fibras pueden estar irregularmente distribuidas, pero también pueden disponerse muy ordenadas: en haces de fibras paralelas, rodeados y separados entre sí, por un conjuntivo de fibras de reticulina.
Se encuentra formando los ligamientos amarillos de los mamíferos. En el hombre además en las cuerdas vocales, el ligamiento estilohiodeo en la laringe, el ligamento suspensor del pene y en los tendones del músculo de la tráquea. También se encuentra en la túnica media de las arterias elásticas.
En la especie humana el tejido adiposo constituye aproximadamente un 10% del peso corporal, y su distribución varía del sexo masculino al femenino. Así en los hombres es abundante en la nuca, en la 7ª vértebra cervical, sobre el deltoides y el tríceps. En mujeres, en los pechos, nalgas y cara anterior del muslo.Se puede considerar como una variante de tejido conjuntivo altamente especializada en el almacenamiento de lípidos. En los mamíferos se encuentra en muchos lugares en donde hay tejido conjuntivo laxo, pero forma acumulaciones mayores en la hipodermis, mesenterios (son distintos repliegues del peritoneo que fijan las distintas porciones del intestino a las paredes abdominales). Este tejido tiene un color blanco-amarillento debido al contenido de carotenoides.
Los adipocitos se encuentran inmersos en una malla de reticulina. Tiene abundantes vasos sanguíneos y es frecuente encontrar células cebadas y leucocitos neutrófilos.
Metabolismo del tejido adiposo
Se basa en el almacenamiento y liberación de triglicéridos según las siguientes etapas:
- Absorción de lípidos en forma de ácidos grasos y glicerol y transporte hasta los adi- pocitos. (ver funciones del epitelio de revestimiento). Son transportados en forma de quilomicrones y sí las grasas han sido sintetizadas en el hígado en forma de lipoproteínas.
- Almacenamiento de los lípidos en los adipocitos en forma de triglicéridos. Para que las lipoproteínas hepáticas y los quilomicrones intestinales puedan penetrar en los adipocitos es necesario que estos compuestos se degraden en monoglicéridos; sólo así los absorbe el adipocito. Esta función la realiza el enzima lipasa lipoproteína, que es sintetizada por los adipocitos y liberada en su proximidad. Una vez absorbidos los monoglicéridos por los adipocitos se convierten en triglicéridos al combinarse con glicerofosfato, y se almacenan en la gota lipídica.
Constituye una variante del tejido adiposo, que se presenta en embriones de mamíferos, siendo más raro en los adultos.
Los adipocitos de grasa parda son células más pequeñas que las del tejido adiposo común. El núcleo no está rechazado pudiendo ser incluso central. El citoplasma no contiene una sola gota de grasa sino muchas gotitas de lípidos de tamaño variable. Se trata de una estructura microvacuolar.
El color de este tejido se debe a la abundancia de citocromos que tienen las mitocondrias de las células de este tejido.
Tipos
|
Características
|
Función
|
Localización
|
Pardo
|
Fi
|
Generación de calor
|
Embriones de mamíferos
|
Blanco
|
Gr
|
Aislante
Reserva de lípidos |
Bajo la piel en homeotermos.
Otras en poiquilotermos |
El adipocito blanco.
El adipocito pardo.
Lámina 8.- Tejido adiposos.
Adiposo: (tráquea)
Es, junto con el hueso, uno de los principales tejidos de soporte de los animales. Su función es posible gracias a las propiedades de su matriz extracelular, la cual es predominante en este tipo de tejido. El cartílago es una estructura semirígida que permite mantener la forma de numerosos órganos, recubre la superficie de los huesos en las artículaciones y es el principal tejido de soporte durante el desarrollo embrionario, cuando el hueso aun no está formado. Es un tejido avascular y su matriz extracelular está formada fundamentalmente por colágeno, fibras elásticas y glucosaminoglicanos sulfatados. Las células que lo componen son los condrocitos, los cuales se localizan en pequeñas oquedades diseminadas denominadas lagunas.
La mayor parte del cartílago, excepto el tipo de cartílago denominado fribrocartílago, está rodeado por una capa de tejido conectivo denominada pericondrio. Éste posee una capa externa, denominada pericondrio fibroso, formada por tejido conectivo fibroso que contiene fibroblastos y fibras de colágeno, y otra interna denominada pericondrio condrogénico, donde se encuentran las células condrogénicas y los condroblastos que por diferenciación darán lugar a los condrocitos.
Las células condrogénicas producen a los condroblastos y estos últimos son los responsables de sintetizar la matriz cartilaginosa. A medida que la sintetizan se van rodeando de ella y se transforman en condrocitos.
El tejido cartilaginoso procede del mesénquima y lo primero que aparece son las fibras colágenas, luego las elásticas, posteriormente las células mesenquimatosas se van a diferenciar y forman los condrocitos. Los condrocitos acumulan vacuolas con lípidos y glucógeno, la masa exterior del cartílago comprime el mesénquima que le rodea y forma una cubierta fibrosa llamada pericondrio.
| Tipos | Características | Función |
Localización
|
Hialino
| Fibras colágenas. Rodeado de fibras elásticas | Resistencia a la presión | Primordios de huesos Esqueleto condictrios. Nariz, externón |
Elástico
| Gran cantidad de fibras elásticas | Flexibilidad | Pabellones auditivos |
Fibroso
| Muchas fibras. Predominantemente colágenas | Resistencia | Discos intervertebrales. |
11.1. Cartílago hialino
Tejido de aspecto cristalino, blanco azulado. Lo encontramos en: caras articulares de los huesos, cartílagos costales, cartílagos de nariz, laringe y traquea, en el feto casi todo el esqueleto es cartílago hialino. Los condrocitos ocupan unas pequeñas cavidades dentro de la matriz rodeados de una laguna.
Formado por células esféricas, nucleo grande y centrado con uno o más nucleolos, citoplasma basófilo, grandes mitocondrias y vacuolas con gotas de agua y glucógeno. En el cartilago vivo los condrocitos ocupan la totalidad de las lagunas, después de un proceso de deshidratación y fijación se produce una cierta concentración de los condrocitos y las células pueden presentar una distorsión notable.
En el centro de un cartílago adulto se pueden encontrar células dispuestas en una misma laguna. A estos grupos se le denomina, nido celular. La matriz o sustancia intercelular puede ser de dos tipos: amorfa o formada. La formada está representada por fibras colágenas.
La matriz es basófila por su contenido en condromucina que es una glucoproteina en el cartílago embrionario tiene disposición homogénea en el adulto irregular alrededor de las células formando las cápsulas del cartílago. El pericondrio tiene células fusiformes no diferenciadas de los fibroblastos, fibras elásticas y colágenas con capilares.
Los condrocitos se nutren de los vasos sanguineos que hay en el pericondrio y de las sustancias alimenticias que atraviesan el tejido.
En las articulaciones rodeando a estas se encuentra la cavidad sinovial que encierra el líquido sinovial, al envejecer, en el cartílago hay menos células, hay cambios regresivos, disminuye la cantidad de condromicina y aumenta la de albúmina. El proceso degenerativo más común del cartílago es la calcificación de la matriz que consiste en depositos de fosfato de calcio en forma de cristales de hidroxiapatito, aumentando el volumen de tejido y produciendose la muerte celular. El cartílago que sufre una lesión se regenera con dificultad, salvo en los niños, en los adultos la regeneración se hace por el pericondrio.



11.2. Cartílago elástico
Básicamente es semejante al cartílago hialino y también está cubierto por pericondrio. Este cartílago no sólo posee fibras de colágeno sino también numerosas fibras elásticas (25 nm de
Los condrocitos se nutren de los vasos sanguineos que hay en el pericondrio y de las sustancias alimenticias que atraviesan el tejido.
En las articulaciones rodeando a estas se encuentra la cavidad sinovial que encierra el líquido sinovial, al envejecer, en el cartílago hay menos células, hay cambios regresivos, disminuye la cantidad de condromicina y aumenta la de albúmina. El proceso degenerativo más común del cartílago es la calcificación de la matriz que consiste en depositos de fosfato de calcio en forma de cristales de hidroxiapatito, aumentando el volumen de tejido y produciendose la muerte celular. El cartílago que sufre una lesión se regenera con dificultad, salvo en los niños, en los adultos la regeneración se hace por el pericondrio.
11.2. Cartílago elástico
Básicamente es semejante al cartílago hialino y también está cubierto por pericondrio. Este cartílago no sólo posee fibras de colágeno sino también numerosas fibras elásticas (25 nm de
Es más elástico que el cartílago hialino y puede soportar mayores deformaciones. También tiene más densidad. Se encuentra en la oreja, la epiglotis, el conducto auditivo externo y formando parte de algunos cartílagos de la laringe
11.2. Cartílago fibroso
Es un carílago intermedio entre el cartílago hialino y el tejido conjuntivo denso alineado. Se caracteriza por su riqueza en fibras de colágeno de diverso grosor, que le confiere gran resis- tencia a las tracciones y compresiones. El colágeno enmascara a toda la sustancia fundamental excepto en la proximidad de los condrocitos.
Esta variedad de cartílago carece de pericondrio.
Se encuentra en los discos intervertebrales, en la sínfisis pélbica, en el menisco y en los puntos en los que los tendones y ligamentos se insertan en el hueso.
El tejido cartilaginosos.
Lamina 11.- Tejido cartilaginoso.
Cartilaginoso Hialino: (tráquea)
12. TEJIDO ÓSEO
El tejido óseo, al igual que el cartilaginoso, es un tejido de sostén en el que la matriz rígida está impregnada por un complejo proteinocálcico que envuelve a las células óseas u osteocitos y sostenida por un armazón de colágeno.
Este tejido cumple varias funciones:
- Una función de sostén, puesto que forma la casi totalidad del esqueleto, y de soporte rígido del organismo en el que se fijan los músculos.
- Una función de protección de órganos vitales. Por ejemplo, el sistema nervioso central está protegido por el cráneo y la columna vertebral, y los pulmones y corazón por la caja torácica.
- Una función hematopoyética, puesto que contiene las células de la médula ósea que generarán las células sanguíneas y linfáticas.
- Actúa como depósito de calcio, fosfato y otros iones, almacenándolos o liberándolos de forma controlada para mantener constante su concentración en los líquidos orgánicos (líquido intersticial, sangre y linfa). Esto posibilita el crecimiento óseo y la reparación de fracturas.
- Un armazón proteico, formado por fibras de colágeno, que representa la mayor parte de su masa.
- Una sustancia fundamental, materia amorfa en apariencia sin estructura, formada por proteoglicanos.
- Una sustancia mineral ósea, insoluble al pH del organismo, que confiere al hueso su dureza y solidez.
- Células òseas, en particular los osteocitos, cada uno de los cuales se encuentra en una cavidad o laguna excavada en la matriz.
Sustancia intercelular mineral: La sustancia intercelular de hueso es dura y presenta unos huecos intercomunicados entre sí en donde se alojan las células óseas.
Las sales minerales del tejido óseo son esencialmente:
- Sales de calcio: fosfato tricálcico, bicarbonato cálcico, fluoruro cálcico , pero sobre todo hidroxiapatito . Estas sales forman como finas agujas que son, en realidad, prismas hexagonales.
- Sales de magnesio y estroncio
- Osteoblastos
- Osteocitos
- Osteoclastos
Osteoblastos: Son las células óseas responsables de la secreción de la sustancia fundamental y de las molÇéculas de colágeno.
Se trata de una célula poliédrica, prismática, provista de numerosas y largas expansiones citoplasmáticas. Su misión es elaborar las fibras de colágeno y la sustancia mineral. Se sitúan en el borde del tejido y son responsables de la osificación (formación de hueso). Esta posición coincide con la capa más interna del periostio.
Son los “formadores de hueso”.
Osteocitos: Son las células óseas de un hueso ya maduro, derivadas de los osteoblastos, y están alojadas en lagunas óseas. Estas células aseguran la renovación y mantenimiento de la matriz ósea y desempeña un papel importante en los intercambios de calcio en la sangre.
Son células alargadas, fusiformes, de 15 a 30 mm de largo por 10 a 15 mm de ancho, que emiten expansiones a través de canalículos, comunicándose unos con otros.

 Osteoclastos: Los osteoclastos son células óseas multinucleadas que desempeñan un papel esencial en la destrucción y reabsorción del hueso.
Osteoclastos: Los osteoclastos son células óseas multinucleadas que desempeñan un papel esencial en la destrucción y reabsorción del hueso.
Se trata de una célula voluminosa de 100 mm de diámetro y que contiene de 10 a 15 núcleos. La zona de la célula en contacto con el hueso tiene abundantes microvellosidades, que recibe el nombre de ribete en cepillo (también presente en otras células).
El osteoclasto contiene numerosos lisosomas cuyo contenido enzimático será el que, una vez expulsado, disuelva el hueso. Estas células actúan en los procesos de reorganización ósea. Se han formado a partir de monocitos.

DISPOSICÓN GENERAL DEL TEJIDO ÓSEO
 Las células y la sustancia intercelular se agrupan formando estructuras típicas llamadas Sistemas de Havers o osteomas. Los sistemas de Havers son sistemas concéntricos de osteocitos en sus lagunas óseas (cavidades en la sustancia intercelular mine- ralizada) alrededor de un conducto, llamado conducto de Havers que contiene un capilar sanguíneo. De este canal parten pequeños canalículos que llegan a las lagunas óseas, comunicando unas con otras. Así se nutren los osteocitos. Los capilares de los canales de Havers se comunican entre sí mediante capilares transversales llamados conductos de Volkman.
Las células y la sustancia intercelular se agrupan formando estructuras típicas llamadas Sistemas de Havers o osteomas. Los sistemas de Havers son sistemas concéntricos de osteocitos en sus lagunas óseas (cavidades en la sustancia intercelular mine- ralizada) alrededor de un conducto, llamado conducto de Havers que contiene un capilar sanguíneo. De este canal parten pequeños canalículos que llegan a las lagunas óseas, comunicando unas con otras. Así se nutren los osteocitos. Los capilares de los canales de Havers se comunican entre sí mediante capilares transversales llamados conductos de Volkman.
Se trata de una célula poliédrica, prismática, provista de numerosas y largas expansiones citoplasmáticas. Su misión es elaborar las fibras de colágeno y la sustancia mineral. Se sitúan en el borde del tejido y son responsables de la osificación (formación de hueso). Esta posición coincide con la capa más interna del periostio.
Son los “formadores de hueso”.
Osteocitos: Son las células óseas de un hueso ya maduro, derivadas de los osteoblastos, y están alojadas en lagunas óseas. Estas células aseguran la renovación y mantenimiento de la matriz ósea y desempeña un papel importante en los intercambios de calcio en la sangre.
Son células alargadas, fusiformes, de 15 a 30 mm de largo por 10 a 15 mm de ancho, que emiten expansiones a través de canalículos, comunicándose unos con otros.
Osteoclastos: Los osteoclastos son células óseas multinucleadas que desempeñan un papel esencial en la destrucción y reabsorción del hueso. Se trata de una célula voluminosa de 100 mm de diámetro y que contiene de 10 a 15 núcleos. La zona de la célula en contacto con el hueso tiene abundantes microvellosidades, que recibe el nombre de ribete en cepillo (también presente en otras células).
El osteoclasto contiene numerosos lisosomas cuyo contenido enzimático será el que, una vez expulsado, disuelva el hueso. Estas células actúan en los procesos de reorganización ósea. Se han formado a partir de monocitos.
DISPOSICÓN GENERAL DEL TEJIDO ÓSEO
Las células y la sustancia intercelular se agrupan formando estructuras típicas llamadas Sistemas de Havers o osteomas. Los sistemas de Havers son sistemas concéntricos de osteocitos en sus lagunas óseas (cavidades en la sustancia intercelular mine- ralizada) alrededor de un conducto, llamado conducto de Havers que contiene un capilar sanguíneo. De este canal parten pequeños canalículos que llegan a las lagunas óseas, comunicando unas con otras. Así se nutren los osteocitos. Los capilares de los canales de Havers se comunican entre sí mediante capilares transversales llamados conductos de Volkman. 12.1. CLASIFICACIÓN DEL TEJIDO ÓSEO
La más sencilla es la macroscópica que se puede establecer a simple vista o con la ayuda de una lupa. Así al examinar la superficie de corte de un hueso seco se puede identificar:
Tejido óseo compacto: de aspecto macizo. En él los Sistemas de Havers se disponen de un modo ordenado y superpuesto.
Tejido óseo esponjoso: presenta múltiples cavidades de tamaño variable como si fuera un esponja. Se observan trabéculas de tejido mineralizado. En el ser humano (y otros vertebrados) contiene la médula Åsea roja. Los sistemas de Havers no son tan cilíndricos sino más amorfos, mucho más grandes y con vasos sanguíneos en disposición más irregular.
El Periostio es una vaina fibrosa que envuelve al hueso, excepto sobre las superficies articulares. Es de naturaleza conectiva y de él parten los vasos sanguíneos que penetran en el hueso por los forámenes y que ramifican sus capilares a través de conductos de Havers y de Volkman.


12. 2. Tipos de huesos y partes de un hueso
Los tejidos óseos se asocian para constituir los huesos, piezas óseas que son verdaderos ór- ganos. Se clasifican en:
Huesos largos: como el fémur, radio, cúbito, ... Los huesos largos se dividen en tres partes:
Diáfisis: o zona larga del hueso. Es cilíndrica y está excavada en su centro por un conducto que contiene médula ósea amarilla. Es de tejido óseo compacto.
Tejido óseo compacto: de aspecto macizo. En él los Sistemas de Havers se disponen de un modo ordenado y superpuesto.
Tejido óseo esponjoso: presenta múltiples cavidades de tamaño variable como si fuera un esponja. Se observan trabéculas de tejido mineralizado. En el ser humano (y otros vertebrados) contiene la médula Åsea roja. Los sistemas de Havers no son tan cilíndricos sino más amorfos, mucho más grandes y con vasos sanguíneos en disposición más irregular.
El Periostio es una vaina fibrosa que envuelve al hueso, excepto sobre las superficies articulares. Es de naturaleza conectiva y de él parten los vasos sanguíneos que penetran en el hueso por los forámenes y que ramifican sus capilares a través de conductos de Havers y de Volkman.
12. 2. Tipos de huesos y partes de un hueso
Los tejidos óseos se asocian para constituir los huesos, piezas óseas que son verdaderos ór- ganos. Se clasifican en:
Huesos largos: como el fémur, radio, cúbito, ... Los huesos largos se dividen en tres partes:
Diáfisis: o zona larga del hueso. Es cilíndrica y está excavada en su centro por un conducto que contiene médula ósea amarilla. Es de tejido óseo compacto.
Epífisis: situadas en los extremos del hueso, están constituidas por tejido óseo esponjoso y por periostio.
Metáfisis: situada entre la diáfisis y la epífisis y separada de ésta última por el cartílago de conjunción.
Metáfisis: situada entre la diáfisis y la epífisis y separada de ésta última por el cartílago de conjunción.
Huesos planos: como el parietal, frontal, omoplato, ... Están cubiertos externamente por tejido óseo compacto y en el interior contienen tejido óseo esponjoso.
Huesos cortos: como los huesos de la falanges. Están formados por tejido óseo esponjoso rodeado de periostio.
El tejido óseo I: las células y sustancia intercelular.
El tejido óseo II: el hueso haversiano.
La osificación endocondral I: el modelo cartilaginoso.
La osificación endocondral II: el manguito óseo.
La osificación endocondral III: la calcificación del cartílago.
La osificación endocondral III: la remodelación.
La articulación sinovial. .
Lámina 12.- Tejido oseo.
Lamina 13.- Un hueso
Compacto: (hueso compacto)
Esponjoso: (hueso esponjoso)
Este tejido, de origen mesenquimático, está constituído por: células musculares (fibras musculares), capaces de generar movimientos al contraerse bajo estímulos adecuados y luego relajarse y tejido conjuntivo estrechamente asociado a las células musculares. Este actúa como sistema de amarre y acopla la tracción de las células musculares para que puedan actuar en conjunto. Además, conduce los vasos sanguíneos y la inervación propia de las fibras musculares.
Tipos de tejido muscular
En los vertebrados, se distinguen 3 tipos de músculo:
- esquelético, estriado o voluntario
- cardíaco, estriado involuntario
- liso, involuntario
Este tejido está formado por manojos de células cilíndricas muy largas (de hasta 30 cm), multinucleadas y estriadas transversalmente, llamadas también fibras musculares esqueléticas. Los núcleos de las fibras se ubican vecinos a la membrana plasmática (sarcolema), que aparece delimitada por una lámina basal (lámina externa). El tejido conjuntivo que rodea a las fibras musculares contiene numerosos vasos sanguíneos y nervios y se dispone de manera de provocar la contracción de las fibras musculares a los sitios de inserción del músculo.
Cada fibra muscular recibe una terminación del axón de una neurona motora, formándose en la zona de unión una estructura denominada placa motora.
El músculo esquelético se une a los huesos a través de los tendones, estructuras continuas con la envoltura conjuntiva llamada epimisio, que rodea externamente al músculo completo. El tejido conjuntivo penetra al interior del músculo formando el perimisio, que corresponde a delgados septos de tejido conjuntivo que envuelven a manojos o fascículos de fibras musculares. A partir del perimisio, se origina el endomisio formado por delgadas vainas de fibras reticulares que rodean cada una de las fibras musculares. Los vasos sanguíneos penetran al músculo a través de estos septos conjuntivos.
La inervación del tejido muscular esquelético se relaciona directamente con la regulación de la contracción de cada fibra muscular y en consecuencia con el estado de tensión del músculo completo.
Fibras musculares esqueléticas
Las fibras musculares esqueléticas se caracterizan por la presencia de estriaciones transversales periódicas. Esta estriación resulta de la existencia en su citoplasma de las miofibrillas estructuras responsables de la contracción muscular.
Las miofibrillas son estructuras cilíndricas largas (1 a 3 mm de diámetro) que corren paralelas al eje logitudinal de la célula, y están formados por miofilamentos finos y miofilamentos gruesos, dispuestos en tal forma que inducen la apariencia de bandas claras y oscuras que se repiten a a lo largo de cada miofibrilla, determinando la organización de los sarcómeros.
La banda oscura se conoce como banda A (de anisótropa) y la clara como banda I (de isótropa). Cada banda I aparece bisectada por una línea transversal oscura denominada disco o línea Z . Al centro de la banda A hay una zona más clara que corresponde a la banda H en cuyo centro está la línea M.
En el citoplasma que rodea a las miofibrillas se disponen tanto las miocondrias como las cisternas del retículo sarcoplásmico a las cuales se asocian los túbulos T, en una organizacion precisa con respecto a los sarcómeros y repetidas a todo lo largo de las células musculares esqueléticas .
Se distinguen 3 tipos de fibras musculares esqueléticas: rojas, blancas e intermedias.
Las fibras rojas, que abundan en los músculos rojos, son de diámetro pequeño y contienen gran cantidad de mioglobina y numerosas mitocondrias, que se disponen en filas entre las miofibrillas y en acúmulos por debajo del sarcolema. Los músculos rojos se contraen más lentamente, por lo que se ha asumido que la fibra roja es una fibra lenta.
Las fibras blancas, presentes en los músculos blancos, son de diámetro mayor, poseen menor cantidad de mioglobina y un número menor de mitocondrias que se disponen, de preferencia, entre las miofibrillas, a nivel de la banda I. En este tipo de fibras la línea Z es mas delgada que en las fibras rojas.
Las fibras intermedias presentan características intermedias entre las otras 2 variedades de fibras, pero superficialmente se asemejan más a las fibras rojas y son mas abundantes en los músculos rojos. Poseen un número de mitocondrias equivalente al de las fibras rojas, pero su línea Z es delgada como en las fibras blancas.
Regulación de la contracción del sarcómero
La contracción muscular está regulada por variaciones en los niveles citosólicos de Ca++, los que afectan las interacciones entre las cabezas de miosina y los filamentos de actina a través de las 2 proteínas accesorias asociadas a actina en el filamento fino: tropomiosina y troponina.
En el músculo en reposo (concentración citosólica de la miosina no puede asociarse a la actina debido a que los sitios de unión para las cabezas de miosina en las G-actina, están bloqueados por la tropomiosina. Al aumentar las concentraciones citosólicas de Ca, la subunidad TnC de la troponina une Ca, produciéndose un cambio conformacional de la molécula de troponina y el desplazamiento de la molécula de tropomiosina hacia la parte mas profunda de la hendidura de la hélice de actina. Como consecuencia los sitios, en la G-actina, capaces de interactuar con las cabezas de la miosina quedan libres.
Las variaciones en las concentraciones citosólicas de Ca se producen en respuesta a los estímulos nerviosos que inducen la contracción muscular y que actúan desencadenando la liberación de Ca desde el retículo sarcoplásmico hacia el citosol.
Estructura de las miofibrillas
La unidad estructural y funcional de las células musculares estriadas es el sarcómero, elemento que se repite a lo largo de las miofibrillas. El análisis de la estructura y composición molecular del sarcómero, permite entender el mecanismo de contracción de las fibras musculares estriadas, basado en el deslizamiento de los miofilamentos gruesos sobre los miofilamentos finos.
Los filamentos gruesos (de 15 nm de ancho y 1.6 mm de largo) están formados principalmente por miosina y se localizan a lo largo de la banda A. Los filamentos finos (de 8 nm de ancho y 1.0 mm de largo) corresponden a microfilamentos de F-actina. Estos anclan en la línea Z, luego cursan a lo largo de la banda I y penetran la banda A, donde corren paralelos a los filamentos gruesos, terminando a nivel de la banda H que contiene sólo filamentos gruesos. En la banda A se observan puentes que se extienden desde los filamentos gruesos hacia los filamentos finos y que corresponden a las cabezas de las moléculas de miosina. A nivel de la línea M cada filamento grueso se asocia a 6 filamentos gruesos adyacentes, a través de puentes proteicos dispuestos radialmente.
Durante el proceso de contracción, los filamentos finos de los sarcómeros adyacentes son empujados hacia el centro de la banda A, lo que produce el acortamiento del sarcómero. Como consecuencia de este proceso, se oblitera la banda H y disminuye la longitud de la banda I, sin que se modifique la longitud de la banda A. El grado de traslapamiento entre filamentos gruesos y finos explica este fenómeno.
Los filamentos finos están formados por actina, tropomiosina y troponinas, proteínas que se relacionan directamente con el proceso de acortamiento del sarcómero .
Los microfilamentos de actina están constituídos por 2 hebras proteicas, que se enrollan para formar una estructura helicoidal doble. Cada hebra corresponde a un polímero de moléculas asimétricas de G actina, lo que otorga a los microfilamentos de actina una polaridad definida.
La molécula de miosina-II está formada por 2 polipéptidos (cadenas pesadas ) y 4 polipéptidos (cadenas livianas). Está organizada en 3 dominios estructural y funcionalmente distintos: cabeza, cuello y cola. En el extremo NH2-terminal las 2 cadenas pesadas presentan una estructura globular, llamada cabeza, la que se continúa en una zona con forma de bastón, de unos 150 nm de largo, cuya porción inicial corresponde al cuello de la molécula y el resto a la cola.
Retículo Sarcoplásmico y túbulos T (túbulos transversos)
En el músculo esquelético, cada miofibrilla está rodeada de un elaborado sistema de membranas lisas que corresponden al retículo sarcoplásmico. Estas membranas están alineadas en forma precisa con respecto al patrón de bandeo de las miofibrillas. En la zona de unión de la banda A con la banda I el retículo sarcoplásmico se expande para formar las cisternas terminales. Las 2 cisternas terminales paralelas se asocian estrechamente a un tubo transverso (T), formando un complejo denominado tríada .
La contracción de una fibra muscular requiere de la contracción simultánea de todas sus miofibrillas. La forma y distribución del sistema T permite que la onda de depolarización, responsable de la contracción muscular, se distribuya rápidamente desde la superficie celular hacia el interior del citoplasma alcanzando a cada miofibrilla.
La depolarización de la membrana plasmática de la célula muscular, que se propaga a lo largo de los túbulos T, produce la apertura de canales de Ca++ en la membrana del retículo sarcoplásmico y la liberación de Ca++ hacia el citosol. Se piensa que la onda de depolarización induce un cambio conformacional en proteínas sensoras del túbulo T, que se transmite directamente a la proteína que forma los canales de Ca++ del retículo sarcoplásmico.
Placa motora (unión neuromuscular)
La membrana plasmática de la célula muscular está electricamente polarizada. Un estímulo apropiado depolariza la membrana y produce la contracción. Normalmente, la depolarización es producida por un estímulo nervioso. El músculo está inervado por terminales nerviosas de neuronas motoras de la médula espinal. En el punto de inervación, el nervio pierde su vaina de mielina, y se asocia a una región especializada de la superficie de la fibra muscular, para formar la placa motora. En una fibra muscular esquelética, cada terminal axónico motor forma sólo una placa motora.
En la zona de contacto, la terminal axónica forma una dilatación que se aloja en una depresión poco profunda de la superficie de la fibra llamada hendidura sináptica primaria .
En la zona de unión, la lámina basal de la célula de Schwann se fusiona con la lámina basal de la célula muscular. Esta cubierta celular fusionada se extiende hacia la hendidura sináptica primaria, separando la fibra nerviosa de la fibra muscular, y penetra al interior de cada hendidura sináptica secundaria. Cuando el potencial de acción alcanza a la placa motora, el neurotransmisor acetilcolina, contenido en las vesículas sinápticas, se libera y difunde a través de la hendidura. Este mediador se une a receptores de acetilcolina presentes en la membrana postsináptica, concentrados principalmente en la entrada de los pliegues sinápticos secundarios, e induce la depolarización del sarcolema que es transmitida a los tubos T.
Unidad motora: conjunto de fibras musculares esqueléticas inervadas por ramificaciones del axón de una misma neurona motora y que, en consecuencia, son estimuladas simultáneamente a contraerse.
Ramas de una misma motoneurona pueden llegar a inervar hasta 500 fibras musculares. Sin embargo, mientras más fino el movimiento que debe efectuar el músculo, menor es el tamaño de la unidad motora, existiendo situaciones en que cada fibra nerviosa inerva sólo una fibra muscular.
El tejido muscular estriado I: organización.
El tejido muscular estriado II.- el citoesqueleto
El tejido muscular estriado III:.el aparato contráctil. la contracción muscular
El tejido muscular estriado IV: La fibra muscular esquelética.
El tejido muscular estriado V: La fibra roja.
El tejido muscular estriado VI: La fibra blanca.
El musculo esquelético: organización.
Lámina 15- Músculo estriado.
Lámina 16- Organización del músculo
Fibra estriada: (esófago, lengua)
El músculo cardíaco está formado por células musculares ramificadas, de estructura estriada y contacción involuntaria, que poseen 1 o 2 núcleos y que se unen entre sí a través de un tipo de unión propia del músculo cardíaco llamada disco intercalar. A diferencia del músculo esquelético, las fibras musculares cardíacas corresponden a un conjunto de células cardíacas unidas entre sí en disposición lineal.
Las células musculares cardíacas, de unos 15 m m de diámetro y unos 100mm de largo, tienen el núcleo ubicado al centro del citoplasma y presentan estriaciones transversales similares a las del músculo esquelético. El retículo sarcoplásmico no es muy desarrollado y se distribuye irregularmente entre las miofibrillas, que no aparecen claramente separadas. Sin embargo, las mitocondrias, que son extremadamente numerosas, están distribuidas regularmente dividiendo a las células cardíacas en miofibrillas aparentes. En el sarcoplasma hay numerosas gotas de lípido y partículas de glicógeno. Las células están rodeadas por una lámina externa, comparable a la lámina basal de los epitelios.
El musculo cardiaco: la fibra de Purkinje.
Cardiaco:(músculo cardiaco)
13.3. Músculo Liso
El músculo liso está formado por fibras musculares lisas que corresponden a células uninucleadas, delgadas y aguzadas en los extremos, cuya longitud varía entre 20 y 500 mm. Este tipo de músculo forma la porción contráctil de la pared de diversos órganos tales como tubo digestivo y vasos sanguíneos, que requieren de una contracción lenta y sostenida. Las células se organizan en grupos, formando haces, rodeados de tejido conjuntivo fibroso que contiene vasos sanguíneos.
El núcleo de las fibras musculares lisas se ubica en el centro de la fibra y los organelos citoplasmáticos tales como mitocondrias, aparato de Golgi, retículo endoplásmico rugoso y ribosomas libres se localizan, mayoritariamente, en la vecindad de los polos nucleares. El resto del citoplasma está ocupado por abundantes miofilamentos finos de actina, una proporción menor de miofilamentos gruesos de miosina, y un citoesqueleto de filamentos intermedios formados por desmina.
Las fibras musculares lisas se disponen desplazadas una respecto de la otra, de manera que el extremo delgado de una fibra se ubica vecino a la parte ancha de la fibra vecina. Esta disposición de las fibras y la localización del núcleo en el centro, explica el aspecto del músculo liso en corte transversal .
Las fibras musculares lisas están rodeadas por una lámina basal (lámina externa) comparable a la lámina basal de los epitelios
El aparato contráctil del músculo liso se contrae mas lentamente que el del músculo estriado, pero permite un acortamiento mayor de las fibras musculares lisas. El mecanismo de contracción, en esta variedad de músculo, también se basa en el deslizamiento de los filamentos finos sobre los filamentos gruesos. Los filamentos de actina de las fibras musculares lisas son fáciles de detectar a nivel ultraestructural; en cambio la visualización de los filamentos gruesos requiere de condiciones de fijación especiales, que demuestran que en el músculo liso por cada filamento grueso hay una proporción mucho mayor de filamentos finos (1:14) que la que se observa en el músculo esquelético (1:6).
En estas células, la contracción es regulada también por alza en las concentraciones citosólicas de Ca++. Sin embargo, la regulación de la contracción está asociada a miosina y no a actina. Un alza en las concentraciones citosólicas de Ca++ induce la fosforilación de las cadenas livianas de la miosina lo que produce una modificación en la cola de la molécula que permite la formación de filamentos gruesos y genera un cambio conformacional en la cabeza que permite su interacción con actina.
Inervación del Músculo Liso
El músculo liso está inervado por nervios de los sistemas simpático y parasimpático. Con frecuencia, los axones de los nervios terminan en una serie de dilataciones en el conjuntivo que rodea a las células musculares. Algunas de estas dilataciones axónicas están muy próximas (10-20 nm) a la superficie de la célula muscular dando origen a uniones neuromusculares. De acuerdo a la proporción de células inervadas en un determinado músculo, se distinguen:
- El tejido muscular liso unitario o visceral, que posee grandes unidades motoras en las que sólo algunas células musculares poseen una unión neuromuscular propia. La excitación se transmite a un número variable de células musculares que no reciben inervación directa, a través de uniones de comunicación (nexos). Esto permite que todas las células musculares de la unidad motora se contraigan o relajen en conjunto.
- El tejido muscular multiunitario presente en órganos que requieren una modulación precisa del grado de contracción de sus células, como el iris del ojo o las arteriolas. En este tipo de músculo liso, las unidades motoras son pequeñas, predominando aquellas en que existe asociación de sólo una célula muscular con cada terminación nerviosa.
La capacidad de regeneración es diferente en las 3 variedades de músculo.
El músculo esquelético tiene la capacidad de regenerar parcialmente a partir de las llamadas células satélite. Estas corresponden a células uninucleadas, fusiformes que yacen dentro de la lámina basal que rodea a cada fibra. Se considera que corresponden a mioblastos que persisten luego de la diferenciación del músculo. Frente a daño muscular u otros estímulos estas células, relativamente escasas, se activan, proliferan y se fusionan para formar nuevas fibras.
El músculo cardíaco no tiene, practicamente, capacidad de regenerar. Los daños del músculo cardíaco se reparan por proliferación del tejido conjuntivo, produciéndose una cicatriz.
El músculo liso tiene tambien una capacidad de regeneración moderada. Luego de daño muscular, algunas células musculares lisas entran en mitosis y reemplazan el tejido dañado. Si la capacidad de proliferación no es suficiente para reparar el daño, se produce una cicatriz de tejido conjuntivo.
ANIMACIONES
14. TEJIDO NERVIOSO
Se origina desde el ectoderma y sus principales componentes son las células, rodeadas de escaso material intercelular. Las células son de dos clases diferentes: neuronas o células nerviosas yneuroglia o células de sostén.
Es el tejido propio del Sistema Nervioso el cuál, mediante la acción coordinada de redes de células nerviosas:
- Recoge información procedente desde receptores sensoriales
- Procesa esta información, proporcionando un sistema de memoria y
- Genera señales apropiadas hacia las células efectoras .
El Sistema Nervioso Central (SNC), se origina desde el epitelio del tubo neural y su tejido nervioso contiene neuronas, células de neuroglia y capilares.
El Sistema Nervioso Periférico (SNP), que conecta los receptores sensoriales con SNC. y a este con las células efectoras, se desarrolla a partir de la cresta neural y sus células se asocian a otros tejidos del organismo. Sin embargo, es una extensión del tejido nervioso del SNC ya que zonas de las neuronas sensitivas y efectoras y todas las interneuronas se encuentran en el SNC, mientras que los ganglios nerviosos y los nervios periféricos corresponde al tejido nervioso propio del SNP
11.1. Neuronas
Son las células funcionales del tejido nervioso. Ellas se interconectan formando redes de comunicación que transmiten señales por zonas definidas del sistema nervioso . Los funciones complejas del sistema nervioso son consecuencia de la interacción entre redes de neuronas, y no el resultado de las características específicas de cada neurona individual.
La forma y estructura de cada neurona se relaciona con su función específica, la que puede se:
- Recibir señales desde receptores sensoriales
- Conducir estas señales como impulsos nerviosos, que consisten en cambios en la polaridad eléctrica a nivel de su membrana celular
- Transmitir las señales a otras neuronas o a células efectoras
- El pericarion que es la zona de la célula donde se ubica el núcleo, y desde el cuál nacen dos tipos de prolongaciones
- Las dendritas que son numerosas y aumentan el área de superficie celular disponible para recibir información desde los terminales axónicos de otras neuronas
- El axón que nace único y conduce el impulso nervioso de esa neurona hacia otras células ramificándose en su porción terminal (telodendrón)
- Uniones celulares especializadas llamadas sinapsis, ubicadas en sitios de vecindad estrecha entre los botones terminales de las ramificaciones del axón y la superficie de otras neuronas
Cada zona de las células nerviosas se localiza de preferencia en zonas especializadas del tejido nervioso:
- Los cuerpos celulares , la mayor parte de las dendritas y la arborización terminal de una alta proporción de los axones se ubican en la sustancia gris del SNC y en los ganglios del SNP
- Los axones forman la parte funcional de las fibras nerviosas y se concentran en los haces de la sustancia blanca del SNC; y en los nervios del SNP
Según el número y la distribución de sus prolongaciones, las neuronas se clasifican en:
Bipolares, que además del axón tienen sólo una dendrita; se las encuentra asociadas a receptores en la retina y en la mucosa olfatoria
Speudo-unipolares, desde las que nace sólo una prolongación que se bifurca y se comporta funcionalmente cono un axón salvo en sus extremos ramificados en que la rama periférica reciben señales y funcionan como dendritas y transmiten el impulso sin que este pase por el soma neuronal; es el caso de las neuronas sensitivas espinales
Multipolares desde las que, además del axón, nacen desde dos a más de mil dendritas lo que les permite recibir terminales axónicos desde múltiples neuronas distintas. La mayoría de las neuronas son de este tipo. Un caso extremo do lo constituye la célula de Purkinje que recibe más de 200.000 terminales nerviosos
- Presentar formas complejas y una gran área de superficie de membrana celular, a nivel de la cuál debe mantener un gradiente electroquímico importante entre el intra y el extracelular
- Secretar distintos tipos de productos a nivel de sus terminales axónicos
- Requerir un recambio contante de sus distintos organelos y componentes moleculares ya que su vida suele ser muy larga (hasta los mismos años que el individuo al que pertenecen).
- El núcleo es grande y rico en eucromatina, con el nucléolo prominente.
- El ergastoplasma que se dispone en agregados de cisternas paralelas entre las cuales hay abundantes poliribosomas. Al microscopio de luz se observan como grumos basófilo o cuerpos de Nissl.
- El aparato de Golgi se dispone en forma perinuclear y da origen a vesículas membranosas, con contenidos diversos, que pueden desplazarse hacia las dendritas o hacia el axón.
- Las mitocondrias son abundantes y se encuentran en el citoplasma de toda la neurona.
- Los lisosomas son numerosos y originan cuerpos residuales cargados de lipofucsina que se acumulan de preferencia en el citoplasma del soma neuronal
- El citoesqueleto aparece, al microscopio de luz, como las neurofibrilla, que corresponden a manojos de neurofilamentos (filamentos intermedios), vecinos a los abundantes microtúbulos
Las principales funciones del axón son:
- El transporte de organulos y moléculas, por el axoplama, entre el pericarion y las ramas del telodendrón.
- La conducción del impulso nervioso, como el desplazamiento del potencial de acción generado por cambios en la permeabilidad a iones a lo largo de la membrana celular axonal (axolema) de las fibras nerviosas
En el SNC los axones están rodeados por la mielina de los oligodendrocitos (fibras nerviosas mielínicas del SNC), mientras que en el SNP pueden estar rodeados, ya sea, por prolongaciones citoplasmáticas de las células de Schwann (fibras amielínicas) o por la mielina las células se Schwann (fibras nerviosas mielínicas del SNP)
Los impulsos nerviosos son ondas transitorias de inversión del voltaje que existe a nivel de la membrana celular, que se inician el sitio en que se aplica el estímulo. Cada una de estas ondas corresponde a un potencial de acción. Este proceso es posible porque entre las macromoléculas que, como proteínas integrales, ocupan todo el espesor del axolema se encuentran:
- La bomba de sodio-potasio, capaz de transportar activamente sodio hacia el medio extracelular intercambiandolo por potasio.
- Canales para Na sensibles a voltaje, que determinan la inversión del voltaje de la membrana ya que al abrirse y permitir la entrada de Na+ hacen que el interior de la membrana se vuelva positiva,
- Canales para K sensibles a voltaje, cuya activación contribuye a la depolarización de la membrana, por salida de iones K desde el interior del axoplasma.
Además, en el interior de la neurona existen proteínas e iones con carga negativa. Esta diferencia de concentración de iones produce también una diferencia de potencial entre el exterior de la membrana y el interior celular. El valor que se alcanza es de unos -70 milivoltios (negativo el interior con respecto al valor de cargas positivas del exterior).
Esta variación entre el exterior y el interior se alcanza por el funcionamiento de la bomba de sodio/potasio (Na+/K+)
La bomba de Na+/K+ gasta ATP. Expulsa tres iones de sodio que se encontraban en el interior de la neurona e introduce dos iones de potasio que se encontraban en el exterior. Los iones sodio no pueden volver a entrar en la neurona, debido a que la membrana es impermeable al sodio. Por ello, la concentración de iones sodio en el exterior es elevada. Además, se pierden 3 cargas positivas cada vez que funciona la bomba de Na+/K+, aunque entren dos cargas de potasio. Esto hace que en el exterior haya más cargas positivas que en el interior, creando una diferencia de potencial. Se dice que la neurona se encuentra en potencial de reposo, dispuesta a recibir un impulso nervioso.
Si la despolarización provoca un cambio de potencial de 120 milivoltios más de los que tenía el interior se dice que se ha alcanzado el potencial de acción, que supone la transmisión del impulso nervioso.
La transmisión del impulso nervioso sigue la Ley del todo o nada. Esto quiere decir que si la despolarización de la membrana no alcanza un potencial mínimo, denominado potencial umbral, no se transmite el impulso nervioso, pero, aunque este potencial sea rebasado en mucho, sólo se envía un impulso nervioso, siempre de la misma intensidad.
Una vez que la neurona emite el impulso nervioso debe volver al inicial potencial de reposo. Para ello, la membrana se repolariza, cerrándose los canales para el sodio que estaban abiertos por la presencia del neurotransmisor. El neurotransmisor es destruido por acción enzimática y el potencial de reposo se alcanza al expulsar el sodio la bomba de Na+/K+.
Sinapsis
Las neuronas, en la mayor parte de los anima les, no se encuentran físicamente unidas. Existe un pequeño espacio entre ellas, llamado hendidura sináptica, al que se vierte el neurotransmisor desde la membrana presináptica, membrana de la neurona que envía el impulso nervioso, a la membrana postsináptica, membrana de la neurona que recibe el impulso nervioso. El neurotransmisor es la molécula responsable de despolarizar la membrana de la neurona que recibe el impulso nervioso, abriendo los canales para el sodio que permanecían cerrados.
En las fibras nerviosas mielínicas, el axón está cubierto por una vaina de mielina formada por la aposición de una serie de capas de membrana celular, que actúa como un aislante eléctrico del axón.A lo largo del axón, la mielina es formadas por células sucesivas y en cada límite intercelular existe un anillo sin mielina que corresponde al nodo de Ranvier.La consecuencia es una conducción saltatoria del potencial de acción ya que la inversión del voltaje inducido a nivel de un nódulo de Ranvier se continúa por propagación pasiva rápida de la corriente por el interior del axón y por el extracelular hasta el nódulo siguiente donde produce la inversión del voltaje. La consecuencia de esta estructura es que en los axones mielínicos la conducción del impulso nervioso es más rápida. La velocidad de conducción del impulso nervioso es proporcional al diámetro del axón y a la distancia entre los nodos de Ranvier.
Uniones intercelulares de las neuronas
Las neuronas se asocian entre sí y con otras células en estructuras especializadas que permiten:
- Inducir la descarga del impulso nervioso de una neurona sensitiva por asociación con células epiteliales de los receptores sensoriales.
- Estimular la acción de células efectoras de otros tejidos, gracias a la capacidad del terminal axónico de asociarse a estas células.
- Transmitir el impulso nervioso desde una neurona a otra mediante estructuras especializadas, las sinapsis, que se encuentran el sitio de contacto de cada terminal axónico con las dendritas o el pericarion de otra neurona.
El tejido nervioso I: tipos de neuronas.
El tejido nervioso II: estructura de la neurona.
El tejido nervioso III: el axón.
El tejido nervioso IV: la mielina en el sistema nervioso periférico.
El tejido nervioso V: la mielina en el sistema nervioso central.
El tejido nervioso VI: estructura de la mielina.
El tejido nervioso VII: la mielinogénesis.
El tejido nervioso VIII: la fibra amielínica.
La sinapsis I: elementos y tipos de sinapsis.
La sinapsis II: histofisiología
La sinapsis III: tipos topográficos y químicos.
La sinapsis IV: la polisinápsis.
La sinapsis V: la sinapsis en barra y bastón.
La sinapsis VI: la sinapsis neuromuscular o placa motora.
Lámina 17- La neurona .
Lámina 18- La membrana y el axon.
Lámina 19- La sinapsis.
Neuronas: (médula, cerebelo, estómago, duodeno)
En el tejido nervioso del SNC, por cada neurona hay entre 10 a 50 células de neuroglia, y que a diferencia de las neuronas retienen su capacidad de proliferar.
Existen 4 clases de células de neuroglia:
- Astrocitos (astroglia)
- Oligodendrocitos (oligodendroglia)
- Células ependimarias
- Microglia
- células de Schwann
- células satélites o capsulares
Tienen formas estrelladas y presentan largas prolongaciones que se extienden hacia las neuronas y hacia los láminas basales que rodean a los capilares sanguíneos (pies terminales), o que separan al tejido nervioso del conjuntivo laxo de la piamadre, constituyendo la glia limitante
Se han identificado dos tipos de astroglia: astrocitos fibrosos que se asocian de preferencia a las fibras nerviosas de la sustancia blanca y astrocitos protoplasmáticos que se concentran de preferencia asociados a los pericariones, dendritas, terminaciones axónicas en la sustancia gris.
Oligodendrocitos
Son más pequeños y con menos prolongaciones que la astroglía.
Su núcleo es rico en heterocromatina y su citoplasma polirribosomas libres, un aparato de Golgi desarrollado y un alto contenido en microtúbulos, tanto en el citoplasma que rodea al núcleo como en sus prolongaciones. Su función más notable es la formación de la mielina, que rodea a los axones del SNC.
El proceso de mielinización del axón por el oligodendrocito es similar al de la célula de Schwann.
Microglia
Se caracterizan por ser pequeñas, con un denso núcleo alargado y prolongaciones largas y ramificadas.
Células de Schwann
Las células de Schawnn se originan de la cresta neural y acompañan a los axones durante su crecimiento, formando la vaina que cubre a todos los axones del SNP desde su segmento inicial hasta sus terminaciones. Ellas son indispensables para la integridad estructural y funcional del axón.
- Fibras nerviosas amielínicas: cuando el axón asociado a la célula de Schwann es de pequeño diámetro se aloja en una concavidad de la superficie de la célula de Schawnn, rodeado por espacio intercelular y conectado hacia el exterior mediante el mesaxón. Varios axones pueden estar alojados de esta forma en la misma célula .
- Fibras nerviosas mielínicas: Los axones de mayor diámetro inducen el proceso de formación de la mielina por la célula de Schwann. En las fibras mielínicas cada célula de Schawnn rodea a solo un axón y su vaina de mielina se ubicada vecina al axón con el resto de su citoplasma en la zona externa. Por fuera, la célula de Schawnn se asocia mediante su lámina basal que al endoneuro.
El citoplasma de la célula de Schwann permanece : junto al axón; junto a la superficie externa de la célula y entre las lamelas internodales de la mielina: en l
El astrocito protoplásmico
El astrocito fibroso.
El oligodendrocito.
La microglia y el ependimocito.
Lámina 20- La glía..
Glía: (médula)
Estructura básica del SNP
En el SNP las células nerviosas se organizan formando dos tipos de estructuras:
- NERVIOS que son haces de fibras nerviosas, mielínicas o amielínicas, mantenidas juntas por tejido conjuntivo organizado en forma específica. Los axones de las fibras nerviosas pueden pertenecer a neuronas motoras del SNC, neuronas sensitivas y neuronas pre o post-ganglionares del SNA
- GANGLIOS que contienen los somas de las neuronas. En los ganglios del sistema nervioso autónomo se encuentran los terminales axónicos de las neuronas pre-ganglionares y el pericarion, dendritas y axones de las neuronas post-ganglionares efectoras.
La unidad estructural es el FASCÍCULO NERVIOSO formado por fibras nerviosas rodeadas por el perineuro, derivado del tejido conjuntivo. El perineuro está formado por células muy aplanadas las cuales se han diferenciado tomando un aspecto epiteliode. El perineuro forma una envoltura continua que acompaña a las fibras nerviosas hasta sus terminaciones ya que juega un rol muy importante en la regulación de la composición del medio que rodea a las fibras nerviosas
Al interior del fascículo las fibras nerviosas están rodeadas por el endoneuro.
Al interior del fascículo las fibras nerviosas están rodeadas por el endoneuro.
Los fascículos nerviosos pueden agruparse formando los NERVIOS propiamente tales, en los cuales varios fascículos se agrupan rodeados por el epineuro que es una vaina de tejido conjuntivo que tiende a ser más bien laxo entre los distintos fascículos pero que se hace bastante fibroso en la periferia, estructurando los troncos nerviosos. El epineuro es un tejido conjuntivo corriente, rico en fibroblastos y en fibrillas de colágeno.
ANIMACIONES
impulso nervioso
CUESTIONES: Nervioso
La sangre se presenta como un líquido complejo que sirve para transportar los gases respiratorios y los principales nutrientes celulares, así como para retirar los productos de deshecho. Llena el sistema cardiovascular y se mueve por un camino cerrado o circuito de vasos, gracias a la acción de bombeo del corazón. La sangre es un medio de transporte complejo que realiza servicios vitales:
- Capta alimento y oxígeno de los aparatos digestivo y respiratorio y los libera a las células, al tiempo que recoge productos de deshecho de las células para llevarlos a los órganos excretores.
- Transporta también hormonas, enzimas, tampones y otras sustancias bioquímicas que cumplen importantes funciones.
- Contribuiye a regular el calor del cuerpo. su elevado calor específico y su conductividad hacen que absorba grandes cantidades de calor, sin que aumente apreciablemente su temperatura, y que lo transfiera desde el centro a la superficie, donde se puede disipar más rápidamente.
15.1. Composición de la sangre
La sangre es más que un simple líquido. No sólo está formada por un líquido, sino también por células y fragmentos de células especializadas denominadas plaquetas. la parte líquida de la sangre, es decir, el plasma es uno de los líquidos principales del cuerpo junto con el intersticial y el intracelular. con el término elementos formes designamos los diversos tipos de células de la sangre y los fragmentos celulares suspendidos en el plasma.
El volumen total de sangre varía considerablemente de unos sujetos a otros. la edad, el tipo corporal, el peso, el sexo y el método de medida son fundamentales. para un hombre adulto joven de 70 kg de peso es de unos 5 litros. Una de las principales variables que modifican el volumen de sangre es la cantidad de grasa corporal. el volumen de sangre por kg de peso corporal varía inversamente con el exceso de grasa corporal. ello significa que cuanta menos grasa se tenga en el cuerpo, más sangre habrá por kilogramo de peso corporal.
15.2. Plasma sanguíneo
El plasma es la parte líquida de la sangre, la sangre completa menos los elementos formes. Para obtener plasma en el laboratorio se centrifuga sangre completa no coagulada. El procedimiento consiste en hacer girar rápidamente la sangre, lo que proyecta las células sanguíneas al fondo de la centrífuga.
El plasma es un líquido claro y de color paja. Contiene un90% de agua y un 10% de solutos. La mayor cantidad de solutos son proteínas 6-8% del plasma. Otros solutos, aunque en cantidades menores son: sustancias alimenticias como glucosa y aminoácidos, sustancias formadas en el meta- bolismo como urea, ácido úrico, CO , sustancias reguladoras como hormonas y otros elementos. Las proteínas presentes en el plasmason de tres clases albúminas (55%), globulinas (38%) y fibrinógeno 7%.
Las proteínas plasmáticas son sustancias de importancia decisiva. Por ejemplo, el fibrinógeno y una albúmina desempeñan un papel clave en la coagulación. las globulinas actúan como elementos esen- ciales en los mecanismos de inmunidad; los anticuerpos circulantes son gammaglobulinas modifica- das. Todas las proteínas del plasma contribuyen al mantenimiento de la viscosidad sanguínea nor- mal, de la presión osmótica de la sangre y del volumen sanguíneo. La síntesis de las proteínas del plasma ocurre en el hígado, excepto de las gammaglobulinas, sintetizadas por células plasmáticas.
15.3. Células sanguíneas
Los elementos formes de la sangre son:
- Glóbulos rojos, eritrocitos o hematies
- Glóbulos blancos o leucocitos
- Plaquetas o trombocitos
Los glóbulos blancos o leucocitos y las plaquetas constituyen menos del 1% del volumen sanguíneo.
GLÓBULOS ROJOS
El glóbulo rojo maduro, normal, no tiene núcleo y su diámetro sólo es de unas 7µm, podría colocarse más de 1500, uno al lado del otro, en el espacio de 1 cm. Antes de que la célula alcance la madurez y penetre en el torrente circulatorio desde la médula ósea, el núcleo es extruido, con la consecuencia de que la célula se hunde por las dos caras. Tienen forma de diminutos discos bicóncavos. el eritrocito maduro es también original al no contener ribosomas, mitocondrias ni otros orgánulos típicos de la mayoría de los cuerpos celulares. Por el contrario, el principal componente de cada glóbulo rojo es el pigmento rojo, llamado hemoglobina. Constituye más de la tercera parte del volumen celular. La depresión de las superficies planas de la célula da lugar a un centro fino con bordes más gruesos. Esta forma original del hematíe le proporciona una superficie muy grande en relación con su volumen. Los bordes de la célula pueden moverse hacia adentro y hacia afuera permitiendo a esta soportar deformidades o cambios de forma sin que la célula se lesione cuando pase por vasos capilares muy estrechos.
Los hematíes son los más numerosos de los elementos formes en la sangre. En el hombre, su número promedio es de unos 5.500.000 por milímetro cúbico de sangre y en la mujer de 4.800.000 por mm3.
Desempeñan un papel fundamental en el transporte de oxígeno y de dióxido de carbono. Ambas funciones dependen de la hemoglobina.
Encerrados dentro de cada glóbulo rojo hay unos 200- 300 millones de moléculas de hemoglobina, que constituyen aproximadamente el 95% del peso seco de cada célula. cada molécula de hemoglobina está formada, como ya sabéis, por cuatro cadenas de proteína. cada una llamada globina está unida a un pigmento rojo (grupo hemo). cada grupo hemo contiene un átomo de hierro. Por tanto una molécula de hemoglobina contiene cuatro átomos de hierro. Este hecho permite a su vez que cada molécula de hemoglobina se una a cuatro moléculas de oxígeno para formar la oxihemoglobina (reacción reversible).
La hemoglobina puede también unirse al dióxido de carbono formando la carboxihemoglobina (también reversible).
En el eritrocito además de hemoglobina, destaca la presencia de un enzima la anhidrasa carbónica, cataliza la reacción que une el dióxido de carbono con el agua, formando ácido carbónico. la disociación del ácido produce iones bicarbonato (HCO -) que se difunde de los hematíes y actúa transportando CO2. en el plasma sanguíneo. Los iones bicarbonato también desempeñan un importante papel en el mantenimiento de los niveles de pH de la sangre.
La sangre del hombre suele contener más hemoglobina que la de la mujer 14- 16 g/100ml frente a 12- 14. Al adulto que no llega a 10 g/100ml se le diagnostica anemia. las anemias se clasifican de acuedo con el tamaño y el contenido de hemoglobina de los glóbulos rojos.
Formación de los hematíes
El proceso completo en la formación de los glóbulos rojos recibe el nombre de eritropoyesis. En el adulto, los eritrocitos comienzan su secuencia de maduración en la médula ósea roja, partiendo de unas células nucleadas llamadas hemocitoblastos o células madres. las células madre tienen capacidad para mantener una población constante de células en diferenciación. se dividen por mitosis y algunas células hijas siguen como células madre indiferenciadas, mientras que otras atraviesan diferentes fases en el desarrollo hasta transformarse en eritrocitos. El proceso de maduración requiere cuatro días.Los glóbulos rojos se forman y destruyen a una velocidad asombrosa 100 millones por minuto. En estado de salud, el número de glóbulos rojos permanece constante, este número es controlado por mecanismos homeostásicos. El ritmo de formación de hematíes se acelera si disminuye la concentración de oxígeno en sangre que llega a los tejidos. La falta de oxígeno eleva las cifras de glóbulos rojos por aumento de secreción de una hormona glucoproteica denominada eritropoyetina. La eritropoyetina estimula a la médula ósea para que acelere la producción de hematíes. la vida media de los glóbulos rojos circulantes es unos 115 - 120 días Muchas veces a medida que envejecen se fragmentan en los capilares. Los macrófagos se encargan de fagocitar a los
hematíes viejos, anormales o fragmentados. El hierro de la hemoglobina es devuelto a la médula ósea para ser reutilizado en la síntesis de nueva hemoglobina. La bilirrubina es transportada al hígado para su excreción al intestino como parte de la bilis. Los aminoácidos liberados son utiliza- dos por el cuerpo bien para obtener energía o para la síntesis de nuevas proteínas. Para formar nuevos hematíes la médula ósea necesita aportes de vitamina B, hierro, aminoácidos, así como cobre y cobalto que actúan de catalizadores.
GLÓBULOS BLANCOS
Hay cinco tipos de glóbulos blancos (GB), o leucocitos, clasificados según la presencia o ausencia de gránulos y las características de tinción de su citoplasma. - Granulocitos: que tienen grandes gránulos en su citoplasma. Se subclasifican en :
- Neutrófilos
- Bsaófilos
- Eosinófilos
- Aganulocitos
- Linfocitos
- Monocito
Neutrófilos: Con colorantes neutros los gránulos de su citoplasma se tiñen de color púrpura muy claro. Estos gránulos son pequeños y numerosos y tienden a dar al citoplasma un aspecto áspero. Se les denomina también leucocitos polimorfonucleares porque sus núcleos tienen dos, tres o más lóbulos
El número aproximado de neutrófilos es del 65% del total de recuentos de GB en una muestra de sangre normal. Estos leucocitos son sumamente móviles y fagocitos muy activos, que pueden emigrar de los vasos y penetrar en los espacios de los tejidos (DIAPÉDESIS). Los gránulos citoplasmáticos contienen potentes lisosomas, que son orgánulos con enzimas de tipo digestivo capaces de destruir las células bacterianas.
Las infecciones bacterianas que provocan una respuesta inflamatoria causan la liberación de sustancias químicas por las células lesionadas, que atraen neutrófilos y otros GB fagocíticos al lugar de la infección. El proceso, llamado quimiotaxis positiva, ayuda al cuerpo a concentrar células fagocíticas en los focos de infección.
Eosinófilos: Contienen numerosos gránulos citoplasmáticos grandes que se tiñen de naranja con los colorantes ácidos como la eosina. Su núcleo suele tener dos lóbulos. Son el 2 - 5% de los GB circulantes.Son numerosos en algunas células del cuerpo como en el recubrimiento de los paratos respiratorio y digestivo. Aunque son fagocitos débiles, pueden ingerir sustancias químicas y proteínas inflamatorias relacionadas con los complejos antígeno- anticuerpo. Sus funciones más importantes tal vez sean de protección frente a las infecciones causadas por gusanos parásitos y a las reacciones alérgicas.
Basófilos : Son relativamente grandes, pero escasos, y sus gránulos citoplasmáticos se tiñen de rojo púrpura oscuro con los colorante básicos. Son los GB menos numerosos, alcanzando sólo el 0,5-1% del recuento total de leucocitos. Son móviles y capaces de diapédesis. Tienen forma de S, pero sus núcleos son borrosos. Los gránulos citoplasmáticos de estos GB contienen histamina (una sustancia química inflamatoria) y heparina (un anticoagulante).
Linfocitos: Son los leucocitos más pequeños, con un diámetro promedio de 6- 8µm. Poseen núcleos grandes, esféricas, rodeados por una cantidad muy pequeña de citoplasma, que se tiñe azul claro. Son con los neutrófilos los GB más numerosos. Constituyen aproximadamente el 25% de toda la población leucocitaria. Dos tipos de linfocitos, denominados linfocitos T y linfocitos B, desempeñan un papel importante en la inmunidad. Los linfocitos T atacan directamente las células infectadas o cancerosas, mientras que los linfocitos B producen anticuerpos frente a antígenos específicos.
Monocitos: Son los leucocitos de mayor tamaño. Sus núcleos son oscuros, tienen forma de habichuela y están rodeados de grandes cantidades de citoplasma de color azul-gris característico. Son células móviles y muy fagocíticas, capaces de absorber grandes bacterias y células infectadas por virus. El número de leucocitos por mm3 es 5000- 9000, con porcentajes distintos de cada clase. Como estas cantidades varían en determinadas situaciones anormales tienen importancia clínica. En la apendicitis aguda, por ejemplo el porcentaje de neutrófilos aumenta, al igual que el recuento total de leucocitos. Denominamos leucopenia a un número excesivamente bajo de GB menos de 5000/ mm3. El SIDA por ejemplo produce una elevada leucopenia. El término leucemia se aplica a un grupo de afecciones malignas caracterizado por la transformación de las células madres que sustituyen a las células normales y provocan leucocitosis (número excesivo de leucocitos) y anemia. Las células leucémicas abandonan la médula ósea y se infiltran por hígado, bazo, SNC, y otras partes del cuerpo.
Formación de leucocitos
La célula madre hemocitoblasto no sólo es precursora de los eritrocitos, sino también de los leucocitos y de las plaquetas de la sangre.
Los granulocitos y algunos linfocitos y monocitos se originan en la médula ósea roja (tejido mieloide). La mayoría de los linfocitos y los monocitos derivan de los hemocitoblastos del tejido linfático. Aunque en la médula ósea se encuentran muchos linfocitos, es probable que la mayoría de ellos se haya formado en tejidos linfáticos y hayan sido transportados a la médula ósea por el torrente circulatorio.
El tejido mieloide y el tejido linfático constituyen los tejidos hematopoyéticos o formadores de células sanguíneas. La médula ósea roja es tejido mieloide en activo, su color rojo proviene de los glóbulos rojos que contiene. La médula ósea amarilla almacena una importante cantidad de grasa, mientras permanece amarilla no es activa en la formación de células sanguíneas, pero puede ser activada cuando se produce una necesidad extrema de hematíes.
PLAQUETAS
Son pequeños cuerpos casi incoloros que por lo general aparecen como discos en forma de huso u ovales que suelen alcanzar 2- 4 µm de diámetro.
Tres propiedades físicas de las plaquetas son: la aglutinación, la adherencia y la agregación. Tan pronto como se extrae sangre de un vaso, las plaquetas se adhieren entre sí y a cualquier superficie con la que entran en contacto; de este modo adoptan formas diversas e irregulares.
El recuento de plaquetas en el adulto arroja un promedio de 250000 por mm3 de sangre. Se consideran normales unos límites de 150.000 – 350000 por mm3. Los niños recién nacidos muestran con frecuencia recuentos más bajos, que aumentan gradualmente para alcanzar los valores normales del adulto hacia los 3 meses de edad. No existen diferencias entre sexos.
Las plaquetas desarrollan una importante función hemostasia y en la formación de coágulos o coagulación. Aunque relacionadas entre sí, ambas funciones son independientes y distintas. Uno a cinco segundos después de la lesión de un capilar, las plaquetas se adhieren al recubrimiento lesionado del vaso y entre sí, formando un tapón hemostático de plaquetas que facilita la detención del paso de sangre a los tejidos. Al menos un tipo de prostaglandinas y ciertas sustancias de origen prostaglandínico, desempeñan un papel en la hemostasia y en la formación de coágulos. Cuando se liberan, estas sustancias actúan sobre el flujo de sangre local (por vasoconstricción) y sobre la agregación plaquetaria en el punto de lesión. Si está es grande, se pone en marcha el mecanismo de formación de coágulos para facilitar la hemostasia.
Las plaquetas se forman en la médula ósea roja, pulmones y, hasta cierto punto, en el bazo por fragmentación de unas células muy grandes (40 – 100 µm), llamadas megacariocitos. estas células se caracterizan por sus grandes núcleos multilobulados que muchas veces tienen forma extraña.
La vida de las plaquetas es corta, unos 9 días por término medio.
15.4. grupos sanguíneos
El término tipo de sangre se refiere al tipo de antígenos (llamados aglutinógenos) presentes en la membrana celular de los hematíes. Los antígenos A, B, y Rh son fundamentales en la sangre en lo referente a las transfusiones y la supervivencia del recién nacido. Se han aislado otros antígenos en la sangre pero su importancia clínica es menor.
SISTEMA ABO
La sangre de todos los seres humanos se corresponde a uno de los cuatro grupos sanguíneos ABO. Estos tipos se denominan de acuerdo con los antígenos presentes en la membrana de los hematíes. He aquí los cuatro tipos ABO:
- Grupo A: antígeno A en los hematíes
- Grupo B: Antígeno B en los hematíes
- Grupo AB: ambas antígenos A y B, en los hematíes
- Grupo O: no hay antígeno A ni B en los hematíes
En la sangre de tipo A el antígeno A está presente en los hematíes, de modo que su plasma no contiene anticuerpos anti- A, pero sí anti- B. En la sangre del grupo B está presente el antígeno B en los hematíes, de manera que el plasma no contiene anticuerpos anti- B, pero sí anti- A. Así pues, la sangre de tipo A donada a un receptor del grupo A no provoca una reacción de aglutinación porque los anticuerpos de tipo B del receptor no se combinan con los antígenos de tipo A de la sangre donada. Sin embargo, la sangre de tipo A donada a un receptor del grupo B provoca una reacción de aglutinación porque los anticuerpos de tipo A del receptor se combinan con los antígenos de tipo A de la sangre donada.
La sangre de tipo O se ha denominado del donante universal, término que significa que se puede administrar con seguridad a cualquier receptor. Sin embargo, esto no es así, ya que el plasma del receptor puede contener aglutininas distintas de los anticuerpo anti-A o anti- B. Por esta razón, la sangre del receptor y del donante cuando son del grupo O deben cruzarse (mezclarse) y observar si existe aglutinación en los hematíes del donante.
La sangre receptora universal (grupo AB) no contiene anticuerpos anti-A ni anti- B, por lo que no puede aglutinar los hematíes de los donantes del tipo a o del tipo B. Ello no significa que cualquier tipo de sangre donada pueda administrarse con seguridad a un individuo con sangre del grupo AB sin cruzarla previamente.
En la sangre del llamado receptor universal, puede haber otras aglutininas que aglutinen antígenos no identificados (aglutinógenos) de la sangre del donante.
SISTEMA Rh
El término Rh positiva significa que está presente el antígeno Rh en sus hematíes. Por otra parte, en la sangre Rh negativa los hematíes no tienen antígeno Rh.
Por lo general ninguna sangre contiene anticuerpos Rh, aunque pueden aparecer en la sangre de las personas Rh negativo si en alguna ocasión han entrado hematíes Rh positivos en el torrente circulatorio. Una forma en que esto puede pasar es administrar una transfusión de Rh + a una persona Rh-. Al cabo de poco tiempo esa persona produce anticuerpos que quedan en la sangre. La otra vía por la que hematíes Rh+ pueden entrar en el torrente circulatorio de un Rh- es durante el embarazo. En ello reside el riesgo de un bebé nacido de una madre Rh- y del padre Rh+. Si el niño hereda ser Rh+ del padre, el factor Rh de los hematíes puede estimular el cuerpo de la madre para que forme anticuerpos anti- R. Por lo tanto si más tarde queda embarazada de otro feto Rh+, este puede padecer una enfermedad llamada eritrobastosis fetal, producida por la reacción entre los anticuerpos Rh de la madre y las células Rh+ del feto. Algunas madres Rh- embarazadas de niños Rh+ son tratadas con una proteína que impide que el cuerpo de la madre forme anticuerpos anti- Rh, evitando así la posibilidad de lesión al siguiente niño Rh+. (eritroblastosis fetal)
Los cromosomas y los genes se localizan por parejas, ya que proceden uno del padre y el otro de la madre. Así, los diferentes genotipos para los distintos grupos sanguíneos son :
-IA IA es la dotación génica responsable del grupo A;
-IA i es la dotación génica híbrida del grupo A;
-IB IB es la dotación génica responsable del grupo B;
-IB i es la dotación génica híbrida del grupo B;
-IA IB es la dotación génica responsable del grupo AB;
-i i es la dotación génica responsable del grupo O.
Lámina 9 .- Celulas inmunocompetentes.
Lámina 10.- Células de la sangre.
Elementos figurados
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Interpretación de un análisis de sangre
Análisis del paciente 1
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Análisis del paciente 3
| ||||||||||||||||||||||||||||||||||||||||||||||||
Análisis del paciente 4
| ||||||||||||||||||||||||||||||||||||||||||||||||
Análisis del paciente 5
| ||||||||||||||||||||||||||||||||||||||||||||||||
Análisis del paciente 6
| ||||||||||||||||||||||||||||||||||||||||||||||||
Las células de la sangre I: los granulocitos.
Las células de la sangre II: el eritrocito
Las células de la sangre III: los agranulocitos.
Las células de la sangre IV: la granulocitopoyesis .
Las células de la sangre V: eritropoyesis.
Las células de la sangre V: plaquetopoyesis.
TEJIDOS ANIMALES
Nombre
|
Ubicación
|
Características
|
Funciones
|
1.- Tejido Epitelial
|
Este tejido incluye la piel y las
membranas que cubren las superficies internas del cuerpo.
|
El epitelio está compuesto por células
estrechamente unidas con escasa sustancia intercelular entre ellas.
- Una de ellas es el epitelio
pavimentoso estratificado presente en la piel y en la superficie del esófago y la vagina.
- Está
formado por una capa fina de células
planas y escamosas que descansan sobre capilares sanguíneos y crecen hacia la superficie, donde mueren y se
eliminan
|
Su principal función es proteger las lesiones e infecciones.
Las principales funciones del tejido
epitelial son las de protección, absorción, secreción y sensación.
|
1.1.- Epitelio de Revestimiento
|
Los tejidos epiteliales limitan tanto las cavidades internas
como las superficies libres del cuerpo.
|
las células se disponen formando láminas.
Presenta una estructura llamada lámina basal.
a) simples, una sola capa
b) estratificados, dos o más capas.
Las formas de las células pueden ser
planas, cúbicas y cilíndricas.
|
Funciona como primera barrera ante agentes patógenos.
|
1.2.- Epitelio Glandular
|
-Las células que conforman el tejido
epiteliar glandular pueden tener forma cilíndrica o cuboide.
-Constituyen aquellas células o asociaciones de células
especializadas para la secreción de sustancia que se encuentran formando las
glándulas que pueden ser uni o pluricelulares.
|
Este tipo de tejido epiteliar tiene
como función secretar sustancias como sudor, leche o cerumen.
|
|
2.- Tejido Nervioso
|
Se ubican en a lo largo del cuerpo
en el sistema nervioso.
|
Cada neurona o célula nerviosa
consta de un cuerpo celular con distintas ramas llamadas dendritas y una
prolongación llamada axón.
Las dendritas conectan unas neuronas
con otras y transmiten información hacia el cuerpo de la neurona; el axón
transmite impulsos a un órgano o tejido.
|
Este complejo grupo de células
transfiere información de una parte del cuerpo a otra; de esta manera
coordina el funcionamiento de un organismo y regula su comportamiento.
|
2.1 .- Neuronas
|
En el sistema nervioso.
|
Es su unidad funcional del sistema nervioso, esta encargada
entre otras cosas es el que transporta el flujo nervioso.
|
|
2.2.- Neuroglias
|
Sistema nervioso
|
Hay alrededor de 10 a 50 veces más células gliales
que neuronas.
Conservan la capacidad mitótica y son las que se encargan de
la reparación (aunque no regeneración) de las lesiones del sistema nervioso.
|
La glía cumple funciones de sostén y nutrición. Esto es
debido a que en el sistema nervioso no existe tejido conjuntivo.
|
3.- Muscular
|
Comprenden los músculos estriados,
lisos y músculos cardiacos.
|
Las células del músculo estriado,
unidas unas con otras, carecen de pared celular y tienen numerosos núcleos y
presentan estrías transversales.
Está constituido por una gran red de
células entrelazadas y vainas musculares.
|
Su función principal es la contracción y relajación de los tejidos.
|
3.1.- Tejido Muscular Liso
|
Se localiza en órganos huecos, excepto corazón, como:
Aparato respiratorio, aparato digestivo, aparato urinario, vasos sanguíneos, etc. |
El músculo liso está formado por fibras musculares lisas que
corresponden a células uninucleadas, delgadas y aguzadas en los extremos,
cuya longitud varía entre 20 y 500 mm
|
Este tipo de tejido opera de manera
independiente a la voluntad del individuo por lo que se conoce a los músculos
que conforma como “músculos involuntarios”.
|
3.2.- Tejido Muscular Estriado
|
Este tipo de tejido es el que
conforma a los músculos que se encuentran unidos a los huesos del cuerpo.
|
Las células
del tejido muscular estriado se caracterizan por tener varios núcleos. Aunque algunos investigadores consideran que la
longitud de las células musculares pueden tener unos
tres centímetros de largo, otros opinan que se
extienden a lo largo de todo el músculo.
Debido a que el músculo formado por tejido muscular estriado se contrae según lo determine el organismo, se le conoce como músculo voluntario.
|
s contracciones del músculo esquelético permiten los
movimientos de los distintos huesos y cartílagos del esqueleto.
|
3.3.- Tejido Muscular Cardiaco
|
El tejido muscular cardiaco
constituye las paredes del corazón.
|
Las células que forman este tejido tienen
bandas transversales microscópicas oscuras y claras que se alternan entre sí.
Los movimientos de este tejido son involuntarios.
Son células alargadas, ramificadas en sus extremos.
|
|
4.- Tejido conectivo o conjuntivo
|
Comprenden el tejido conectivo
elástico y fibroso, el tejido adiposo (tejido graso), el cartílago y el hueso
|
A diferencia del epitelio, las
células de estos tejidos están muy separadas unas de otras, con gran cantidad
de sustancia intercelular entre ellas.
La sangre y la linfa suelen
considerarse tejidos conectivos.
|
Estos tejidos, en conjunto, sustentan y mantienen las
distintas partes del cuerpo
|
4.1.- Tejido adiposo
|
Se encuentran entre el citoplásmico periférico y el núcleo
excéntrico
|
El tejido adiposo es uno de los tejidos más abundantes y
representa alrededor del 15-20% del peso corporal del hombre y del 20-25% del
peso corporal en mujeres.
Los adipocitos diferenciados pierden la capacidad de
dividirse; sin embargo, son células de una vida media muy larga y con
capacidad de aumentar la cantidad de lípidos acumulados.
|
Cumple funciones mecánicas: una de ellas es servir como
amortiguador, protegiendo y manteniendo en su lugar los órganos internos así
como a otras estructuras más externas del cuerpo, y también tiene funciones
metabólicas.
|
4.2.-Tejido cartilaginoso
|
El tejido cartilaginoso se ubica en
ciertas posiciones del organismo, por ejemplo, en las articulaciones.
|
Este tejido presenta células
estrechamente unidas y poco material intercelular, pero a diferencia del
tejido óseo presenta gran flexibilidad, sin dejar de ser muy resistente.
Los animales en su etapa embrionaria
no tienen huesos, en lugar de eso, el embrión mantiene su forma gracias a un
esqueleto formado por cartílago.
|
Sirve de unión entre huesos y
músculos, etc.
|
4.3 .-Tejido óseo
|
Se encuentran en los huesos.
|
Este tipo de tejido se caracteriza
por presentar células muy unidas y con poca materia intercelular. Las
estructuras así formadas suelen ser muy sólidas y resistentes.
Las células del tejido óseo son las
que forman los huesos, por lo que se encuentran distribuidas en el esqueleto
animal.
|
Entre las funciones del tejido óseo
se encuentra la de sostener el resto del organismo, la de darle forma, la de
proteger a los órganos internos y la de colaborar con los movimientos.
|
4. 4 .-Tejido hematopoyético
|
El tejido hematopoyético junto con el tejido adiposo, son
los principales componentes tisulares de la médula ósea.
|
Es la producción de las células de la sangre mediante un
proceso llamado hematopoyesis
|
|
4.5.-Tejido sanguíneo
|
El tejido sanguíneo se encuentra
distribuido a través de todo el organismo.
|
El tejido sanguíneo está compuesto
por los glóbulos rojos (eritrocitos), los glóbulos blancos (leucocitos:
linfocitos, monocitos, neutrófilos, eosinófilos, y basófilos) y las plaquetas
(trombocitos). Además, estas células se encuentran suspendidas en una
sustancia llamada plasma sanguíneo.
|
Las funciones de este tipo de tejido
son las de transporte de sustancias, la de defensa del organismo y participar
en la reparación del organismo.
|
Tejidos animales
17. REPASO


Tejidos animales
Test tejidos animales 1
Test tejidos animales 2
Test tejidos animales 3
Test tejidos animales 4
Tejidos animales 5
Tejidos animales 6
CUESTIONES: 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 1 2 3 26 42 43 44 45 46 47
18. PRÁCTICAS
Tejidos animales
Test tejidos animales 1
Test tejidos animales 2
Test tejidos animales 3
Test tejidos animales 4
Tejidos animales 5
Tejidos animales 6
18. PRÁCTICAS
Microscopio y epitelio cebolla
Histología animal 1
Histología animal 2
Histología animal 3
Sangre
19. OTRAS PRESENTACIONES
Historia del microscopio
Organización pluricelular
Tejidos animales y vegetales
Tejidos animales y vegetales
Tejidos animales
Tejidos animales 1
Tejidos animales 3
Tejidos animales 5
Tejidos animales 6
Tejidos animales 7
Epitelio
Tejidos epiteliales
Epitelio glandular
Tejido conjuntivo
Epitelio de revestimiento
Cartilagos del cuerpo
Tejido muscular
Tejido nervioso
Tejido nervioso
Células gliales
Histología animal 1
Histología animal 2
Histología animal 3
Sangre
19. OTRAS PRESENTACIONES
Historia del microscopio
Organización pluricelular
Tejidos animales y vegetales
Tejidos animales y vegetales
Tejidos animales
Tejidos animales 1
Tejidos animales 3
Tejidos animales 5
Tejidos animales 6
Tejidos animales 7
Epitelio
Tejidos epiteliales
Epitelio glandular
Tejido conjuntivo
Epitelio de revestimiento
Cartilagos del cuerpo
Tejido muscular
Tejido nervioso
Tejido nervioso
Células gliales
20. OTRAS IMÁGENES
1. SISTEMA LINFOIDE: Bazo, Ganglio linfático, Timo
2. SISTEMA ENDOCRINO: Suprarrenal, Tiroides
3. SISTEMA RESPIRATORIO: Tráquea, Parénquima pulmonar
5. SISTEMA REPRODUCTOR FEMENINO: Pared uterina, Ovario y trompa, Glándula mamaria
6. SISTEMA REPRODUCTOR MASCULINO: Testículo y epidídimo, Glándula prostática
7. SISTEMA DIGESTIVO: Esófago y estómago, Intestino delgado y grueso, Vesícula biliar / Pancreas / Hígado16.
Cuestiones histología 2
Cuestiones histología 3
Cuestiones histología 4
Cuestiones histología 5
Cuestiones histología 6
Cuestiones histología 7
Preguntas con imágemes
Preguntas variadas
Test histología 1
Test histología 2
Test histología 3
Test histología 4
Test histología 5
Cuestiones histología 3
Cuestiones histología 4
Cuestiones histología 5
Cuestiones histología 6
Cuestiones histología 7
Preguntas con imágemes
Preguntas variadas
Test histología 1
Test histología 2
Test histología 3
Test histología 4
Test histología 5